Sistemas Sensoriais/Sistema Visual
Introdução
editarDe uma forma geral, o sistema visual depende de ondas electromagnéticas (EM) de forma a dar a um organismo mais informações sobre o meio que o rodeia. Esta informação pode ser muito variada e pode envolver possíveis parceiros para acasalamento, perigos e fontes de subsistência. Os vários organismos desenvolveram constituintes diferentes para o que apelidamos de sistema visual.
A complexidade de um olho pode ir de uma pequena colecção de células fotorreceptoras a um órgão complexo tipo câmara fotográfica. Se um organismo possuir diferentes tipos de células sensíveis à luz, ou fotorreceptores sensíveis a diferentes tipos de comprimentos de onda, então é em teoria capaz de perceber cores, ou pelo menos distinguir cores díspares. Alguns organismos, em particular certos insectos e cefalópodes, são também capazes de detectar a polarização da luz.
É importante aqui referir que o foco deste texto serão as ondas EM. É verdade que certos organismos desenvolveram formas alternativas de "visão", ou pelo menos métodos que lhes permitem complementar o que vêem com informação extra-sensorial. Um exemplo óbvio será a ecolocalização usada por baleias e morcegos. No entanto, o uso de palavras como "visão" e "visual" neste contexto não estão correctos. Estes termos costumam ser reservados para quando falamos de ondas EM dentro do espectro visível, cujos limites costumam ser associados aos do espectro visível humano. Contudo, visto que muitos organismos detectam frequências acima e abaixo dos limites humanos, dever-se-ia apostar numa melhor definição. Assim, definimos aqui o espectro visível como o intervalo de ondas EM que vai dos 300 aos 800 nm. Esta definição poderá parecer arbitrária, mas a verdade é que seleccionar limites errados implicaria considerar certos aspectos da visão de pássaros como não-visão. Por outro lado, neste intervalo não é incluído a visão térmica de alguns organismos, como a de serpentes. Ou seja, serpentes que usem as suas fossetas loreais (sensíveis a ondas EM entre os 5000 e os 30 000 nm) não "vêem", mas sim "sentem".
Neste capítulo será primeiro feita uma breve descrição dos vários tipos de órgãos visuais, seguida de uma explicação aprofundada dos componentes da visão humana, do processamento de sinal das vias visuais nos humanos. Por fim, a percepção em si será analisada.
Órgãos sensoriais
editarA visão, ou a capacidade de ver, depende dos órgãos sensoriais disponíveis. Existem várias variantes do que nós chamamos "olhos", com níveis de complexidade muito diferentes. As várias variantes oferecem diferentes vantagens e possibilidades, são sensíveis a comprimentos de onda diferentes e possuem diferentes níveis de acuidade visual. Por outro lado, o processamento de sinal também será dependente das necessidades de cada olho e o número de olhos necessários para uma visão adequada também varia. A capacidade de detectar e decifrar ondas EM provou ser vantajoso para muitos organismos, oferecendo uma maior capacidade de sobrevivência. Em ambiente com pouca ou sem luz, um sistema visual não traz nenhum benefício adicional, o que resultou em atrofia deste sistema em muitos animais (e.g. animais que vivem em grutas, morcegos), com desenvolvimento subsequente de outros aparelhos sensoriais. É interessante notar que os órgãos visuais parecem ajustados à janela óptica, isto é, aos comprimentos de onda (300 a 1100 nm) que atravessam a atmosfera e atingem o solo terrestre. Ao consultar a figura abaixo, poderá notar a existência de outras "janelas", como a janela de infravermelhos (o que explicará até certo ponto a "visão" térmica das serpentes) e a janela de radiofrequências, que nenhum ser vivo conhecido consegue detectar.

Ao longo do tempo, a evolução tem nos apresentado muitas possíveis variantes de olhos, sendo que alguns evoluíram várias vezes, dando origem a semelhanças entre ser vivos de nichos semelhantes. Contudo, existe um aspecto comum a todas as espécies, independentemente da complexidade do órgão sensorial: o uso de opsinas. Sem querer colocar um foco demasiado grande na base molecular, as várias variantes de órgãos sensoriais podem ser divididas nos seguintes grupos:
- Machas oculares
- Stemmata
- Olhos pinhole
- Olhos com cristalinos
- Olhos com córnea refractiva
- Olhos reflectores
- Olhos compostos
As configurações menos complexas de olhos permitem uma simples detecção da luz ambiente, informando o organismo da existência ou ausência de luz. Nestes casos, o aparelho sensorial é normalmente constituído por uma simples concentração de células fotorreceptoras, donde o nome "mancha ocular". Ao adicionar estruturas angulares e recessos, os organismos ganham acesso a informação direccional, o que constitui uma condição crucial para a formação de imagens.
As stemmata são de longe o tipo de aparelhos sensoriais visuais mais comuns e podem encontradas em 95% de todas as espécies.
.jpg)
Levando esta estratégia a um extremo, a fosseta pode ser transformada numa estrutura cavernosa, o que aumenta a acuidade da imagem, mas infelizmente à custa de uma perca de intensidade. Dito de outra forma, existe um trade-off entre intensidade e acuidade. O exemplo a considerar será o Nautilus, espécie da família Nautilidae, organismos considerados fósseis vivos; esta espécie é a única conhecida que possui um olho pinhole, que é completamente análogo à câmara pinhole ou à câmara obscura. Por outro lado, e tal como câmaras fotográficas mais avançadas, os Nautili conseguem ajustar o tamanho da fenda, conseguindo regular a resolução e a luminosidade da imagem. Tal como na máquina fotográfica, uma forma de aliviar o problema do trade-off entre intensidade e resolução e adicionando um cristalino, uma lente, que concentra a luz numa estrutura central, que muita vezes contém uma concentração maior de fotorreceptores. A capacidade de ajustar a forma da lente e de a movimentar, aliada à possibilidade de controlar o tamanho da fenda (ou pupila), deu a organismos a aptidão de se adaptarem a diferentes condições e de focarem em diferentes regiões de interesse da cena visual. O último grande avanço nas variantes de olhos foi a adição de uma córnea refractiva. Olhos com esta estrutura delegam dois terços do seu potência óptica total ao alto índice refractivo da córnea, o que permite uma visão de alta resolução. Muitos animais terrestres, incluindo humanos, possuem esta peça chave. Por outro lado, muitas variantes na estrutura e número de cristalinos, na densidade dos fotorreceptores, no tamanho e número de fóveas, na forma da pupila etc. existem, com o intuito de aumentar a capacidade de sobrevivência do organismo em questão. Toda esta diversidade leva a uma aparência muito variada de olhos, mesmo dentro da variante de olhos únicos. Para demonstrar este ponto, uma colecção de olhos do mesmo tipo (com córnea refractiva), foi compilada na imagem abaixo:
 |
 |
 |
 |
.jpg) |
Uma alternativa à construção de um cristalino serão os olhos reflectores, presentes por exemplo em moluscos. em vez de focarem a luz na porção posterior do olho usando uma lente ou um sistema de lentes, estes organismos possuem estruturas tipo espelho no interior da câmara do olho que reflectem a luz para uma porção central (no fundo como um reflector parabólico). Embora não existam exemplos conhecidos de organismos com olhos reflectores capazes de formar imagens, pelo menos uma espécie de peixes (Dolichopteryx longipes) usa este tipo de olho em combinação com olhos com cristalino.

O último grupo de olhos, os olhos compostos, são encontrados em insectos e crustáceos. Estes olhos são compostos por subunidades funcionais denominadas omatídeos. Cada omatídeo é composto por uma faceta, um cristalino transparente e células fotorreceptoras. É de referir que os omatídeos se encontram separados por células pigmentadas, garantindo que a luz entra o olho o mais paralela possível. A combinação do output do todos os omatídeos forma uma imagem em mosaico, com resolução proporcional ao número de omatídeos. Se os humanos tivessem olhos compostos e quisessem manter a mesma resolução visual, estes teriam coberto toda a nossa face. Como nota, existem vários tipos de olhos compostos, mas não nos compete aqui entrar em grande detalhe nesse subtema.
Explorámos ainda agora muitas variantes de olhos, mas é preciso saber que também o número de olhos varia. Como saberão, os humanos possuem comummente dois olhos, enquanto, em aranhas, este número varia (embora a maioria das espécies possua oito). Também é usual, em aranhas, os diferentes pares de olhos variarem em termos de tamanho, sendo cada tamanho associado a uma função diferente. Por exemplo, as aranhas saltadoras possuem dois grandes olhos frontais que lhes oferecem uma excelente acuidade visual, útil para caçarem as suas presas. Os seus outros seis olhos têm uma resolução muito inferior, mas ajudam as aranhas a evitar predadores. Duas fotografias dos olhos de uma aranha-saltadora e de uma aranha-lobo são expostas para demonstrar a variabilidade topológica ocular dos aracnídeos.
- Topologia ocular das aranhas
-
 Aranha-lobo
Aranha-lobo -
 Aranha-saltadora
Aranha-saltadora


Anatomia do sistema visual
editarNós humanos somos criaturas visuais e por isso mesmo observamos uma grande complexidade em muitos componentes dos nossos olhos. Neste capítulo, iremos tentar descrever estes componentes, de forma a transmitir algum conhecimento sobre as propriedades e funcionalidades da visão humana.
We humans are visual creatures, therefore our eyes are complicated with many components. In this chapter, an attempt is made to describe these components, thus giving some insight into the properties and functionality of human vision.
Entrando no olho - Pupila, íris e cristalino
editarOs raios de luz entram no nosso olho através de uma fenda (a pupila), presente na porção anterior do olho. O aspecto preto é devido ao facto da luz ser completamente absorvida pelo tecido no interior do olho. A luz apenas consegue entrar no olho através da pupila, o que significa que a quantidade de luz que é detectada é determinada pelo diâmetro pupilar. Um esfíncter pigmentado, a íris, rodeia a pupila e é responsável pelas diferentes "cores de olhos" presentes em humanos.
Para além desta camada de pogmentos, a íris tem duas camadas de músculos ciliares. O esfíncter pupilar forma um músculo circular que ajuda a diminuir o diâmetro pupilar e constitui a primeira camada muscular. A segunda camada contém um músculo dilatador da pupila. Estes dois músculos conseguem portanto dilatar ou contrair a pupila em função do meio ambiente e do estado vegetativo da pessoa. Os músculos ciliares estão também em contacto com as fibras zonulares, que também permitem alterar a forma do cristalino e mantê-lo na sua posição anatómica.
O cristalino está situado imediatamente atrás da pupila. A sua forma e características revelam uma função semelhante à das lentes de câmaras fotográficas. No entanto, o cristalino funciona de forma algo diferente. A conformação do cristalino é ajustada pela tracção exercida pelas fibras zonulares, que assim controlam a distância focal. Juntamente com a córnea, o cristalino também permite alterar o foco, o que faz dele uma estrutura de facto muito importante. No entanto, apenas um terço da potência óptica é devido ao cristalino. O cristalino também serve de filtro. As fibras do cristalino são o seu maior constituinte e são compostas de longas células anucleadas despidas da maior da maquinaria celular habitual. O objectivo aqui é garantir a transparência desta lente. Estas fibras, juntamente com proteínas hidrossolúveis chamadas cristalinas, aumentam o índice refractivo do cristalino. As fibras do cristalino também desempenham um papel na manutenção da estrutura e forma do cristalino.

A córnea e a esclera
editar
A córnea, que é responsável pelos restantes dois terços da potência ocular do olho, cobre a íris, a pupila e o cristalino. A córnea foca os raios de luz que irão passar pela íris e pelo cristalino. Tem uma espessura de apenas 0.5 mm e é constituída por cinco camadas:
- Epitélio: uma camada de tecido epitelial que recobre a superfície da córnea.
- Camada de Bowman: uma camada protectora espessa e composta de fortes fibras de colagénio que ajudam a manter a forma da córnea.
- Estroma: uma camada composta de fibras de colagénio paralelas. Esta camada constitui cerca de 90% da espessura da córnea.
- Membrana de Descemet e endotélio: duas camadas adjacentes à câmara anterior do olho, repleta do humor aquoso produzido pelo corpo ciliar. O humor aquoso ajuda a hidratar o cristalino, limpa-o e contribui para a manutenção da pressão ocular. A câmara anterior, situada entre a córnea e a íris, contém um corpo trabecular através do qual o humor aquoso é drenado para o canal de Schlemm e deste para a rede venosa episcleral.
A superfície da córnea situa-se por baixo de duas camadas protectoras: a esclera e a cápsula de Ténon. Ambas estas camadas protectoras envolvem o globo ocular. A esclera é feita de colagénio e fibras elásticas que protegem o olho de lesões externas e são responsáveis pela cor branca do olho. A esclera é atravessada por nervos e vasos e apresenta-se posteriormente em continuação com as meninges que recobrem o nervo óptico. Por outro lado, é recoberta pela conjuntiva, que constitui uma membrana mucosa transparente que também recobre a porção interior da pálpebra. É coberta pela lágrima produzida pela glândula lacrimal. As pálpebras também desempenham uma função protectora e ajudam a espalhar a lágrima na superfície do olho.
Movimentando os olhos - os músculos extra-oculares
editarO globo ocular é movimentado por um conjunto complexo de músculo extra-oculares constituído por quatro músculos rectos (inferior, superior, medial e lateral) e dois oblíquos (superior e inferior).

Como é possível ver, os músculos extra-oculares (2,3,4,5,6,8) possuem uma inserção na esclera e originam-se no tendão de Zinn, um tendão fibroso que rodeia o nervo óptico. Um sistema de polia é criado com a tróclea enquanto polia e o músculo oblíquo superior actuando como corda (esta construção é necessária para dirigir a força muscular de forma correcta). Os músculos extra-oculares restantes não formam este sistema de polia. Usando estes músculos extra-oculares, o olho pode ser rodado para cima, baixo, para a esquerda, para a direita e vários outros movimentos que resultem das combinações destes primeiros podem ser efectuados.
Existem movimentos muito importantes para a nossa visão. Por exemplo, movimentos de vergência permitem um bom funcionamento da visão binocular. Movimentos rápidos e inconscientes chamados sacadas são essenciais para manter a visão fixada num objecto. Uma sacada é uma espécie de movimento agitado que é executado quando os olhos estão a inspeccionar o campo visual e que permite alterar discretamente o ponto de fixação. Quando segue um objecto em movimento, os seus olhos fazem chamado movimentos de perseguição, que se pretendem contínuos. Movimentos involuntários como o nistagmo são causados por sinais oriundos do sistema vestibular e integram o reflexo vestíbulo-ocular.
O tronco encefálico controla todos os movimentos oculares, com diferentes áreas responsáveis por diferentes movimentos.
- Ponte: movimentos rápidos horizontais (por exemplo: sacadas e nistagmo)
- Mesencéfalo: movimentos verticais e de torsão
- Cerebelo: afinamento dos movimentos
- Núcleo de Edinger-Westphal: movimentos de vergência
Onde a detecção da luz ocorre - a retina
editar
Antes de haver transdução, as ondas EM têm de passar pela córnea, pelo cristalino e pela mácula. Estas estruturas também funcionam como filtros que ajudam a diminuir a quantidade de ondas EM indesejáveis, protegendo assim o olho de radiações possivelmente nocivas. Na figura acima, é possível observar que a córnea atenua os comprimentos de onda menores, afectando apenas minimamente cumprimentos de onda maiores. O cristalino bloqueia cerca de 25% das ondas EM abaixo dos 400 nm e mais de 50% das ondas abaixo dos 430 nm. Finalmente, o epitélio pigmentado, que representa o último filtro antes de haver fotorrecepção, afecta cerca de 30% das ondas entre os 430 e os 500 nm.
A parte do olho que marca a fronteira entre a porção fotossensível e não-fotossensível do olho é chamada a ora serrata. A porção sensível à luz do olho é denominada retina e constitui o elemento sensorial de todo este aparato. A retina é composta de múltiplas camadas e contém milhões de fotorreceptores: os cones e os bastonetes, que capturam os raios de luz e iniciam o processo que os irá transformar em impulsos eléctricos. A transmissão desses impulsos é iniciada nas células ganglionares e feita através do nervo óptico.

À direita encontra-se uma ilustração conceptual das estruturas da retina. Como pode ser observado, existem cinco principais tipos de células:
- células fotorreceptoras
- células horizontais
- células bipolares
- células amácrinas
- células ganglionares
As células fotorreceptoras podem por sua vez ser divididas em dois subtipos: os cones e os bastonetes. Os cones encontram-se em menor número na maioria da retina, mas formam um enorme agregado na mácula, especialmente na porção central da fóvea. Nesta região central, cada cone está conectado a uma única célula ganglionar. Por outro lado, os cones desta região são ligeiramente mais pequenos que o cone médio, aumentando a sua concentração na fóvea. É exactamente por causa deste rácio cone/célula ganglionar e da alta concentração de cones na fóvea que é nesta região que apresentamos a maior acuidade visual.

O olho humano possui três tipos de cones, cada cone respondendo a um intervalo específico de comprimentos de onda. Isto é devido à presença de três variantes de um pigmento chamado fotopsina. Visto que os pigmentos são sensíveis a luz vermelha, azul ou verde, assim dizemos que temos cones "vermelhos", "azuis" ou "verdes", também denominados cones S (de short wavelength), M (de medium wavelength) ou L (long wavelength), respectivamente. A fotopsina é constituída por uma proteína chamada opsina e um cromóforo chamado retinal. As partes constituintes de um cone são: o terminal sináptico, os segmentos interiores e exteriores e os seus organelos celulares, em particular o núcleo e as mitocôndrias.
As sensibilidades espectrais dos três tipos de cones:
- Os cones de tipo S absorvem luz com um comprimento de onda curto, isto é, luz azul-violeta. A absorção máxima dá-se com o comprimento de onda 420 nm.
- os cones de tipo M absorvem luz azul-verde a amarela. Neste caso, o comprimento de onda de 535 nm é que está associado à máxima absorção.
- os cones de tipo L absorvem luz amarela a vermelha. O comprimento de onda de maior absorção é 565 nm.

O segmento interno de um cone contém o núcleo celular e os seus organelos. O pigmento transmembranário está localizado no segmento externo, dentro das invaginações da membrana (que formam aparentes discos claramente visíveis na figura à direita). Estes discos maximizam a área receptiva das células. Os cones de muitos vertebrados contêm organelos esféricos (descritos como sendo parecidos com gotículas de óleo), que poderão constituir filtros intra-celulares cuja função será aumentar o contraste, diminuir o brilho e atenuar certas aberrações cromáticas causadas pelo gradiente de tamanho mitocondrial da periferia ao centro celular.
Os bastonetes têm uma estrutura similar à dos cones, mas contêm um pigmento diferente: a rodopsina. Este pigmento permite-lhes detectar luz de fraca intensidade and torna os bastonetes 100 vezes mais sensíveis que os cones. A rodopsina é o único pigmento encontrado nos bastonetes humanos e encontra-se, à semelhança dos cones, no segmento externo dos bastonetes, contido nas invaginações membranares que formam, mais uma vez, um aparente disco. Para além disso, os bastonetes, tal como os cones, também se ligam através do terminal sináptico a uma célula bipolar e os segmentos interno e externo encontram-se ligados através de um cílio conectante.
A rodopsina absorve luz entre os 400 e os 600 nm, apresentando uma absorção máxima cerca dos 500 nm. Este comprimento de onda corresponde a luz azul, o que explica a razão pela qual a cor azul parece mais intensa em relação a, por exemplo, a cor vermelha, à noite.

Ondas EM com comprimentos de onda fora do intervalo 400-700 nm não são detectados nem por cones, nem por bastonetes, o que significa que não são visíveis para os seres humanos.
As células horizontais ocupam a camada nuclear interna da retina. Existem dois tipos (A e B) de células horizontais e ambos respondem à luz hiperpolarisando, isto é, diminuindo o seu potencial de membrana. As células de tipo A incluem um subtipo HII-H2 que interage primariamente com cones S. As de tipo B contêm um subtipo chamado HI-H1, com uma árvore dendrítica e um axónio. O primeiro contacta sobretudo com cones M e L e o segundo com bastonetes. O contacto com cones é maioritariamente feito através de sinapses inibitórias, enquanto que as células estão unidas através de uma rede de gap junctions.

As células bipolares estendem processos dendríticos únicos para a membrana plexiforme externa e os seus corpos celulares encontram-se na membrana nuclear interna. Os processos dendríticos conectam-se exclusivamente a cones e bastonetes e existem 10-11 tipos diferentes de células bipolares (um tipo que se conecta a bastonetes e 9 a 10 outras variantes que liga a cones). As células bipolares ligam-se através do terminal axónico às células amácrinas e ganglionares da camada plexiforme interna. As células bipolares que se conectam a bastonetes ligam-se a tríades sinápticas ou a 18 a 70 bastonetes. Os seus axónios ligam-se aos terminais sinápticos da camada plexiforme interna, que contém ribbon synapses e contacta com processos celulares formando díades sinápticas. Estas células bipolares encontram-se conectadas a células ganglionares com ligações a células amácrinas do tipo AII.
As células amácrinas podem ser vistas na camada nuclear interna e na camada de células ganglionares da retina. Ocasionalmente também são observadas na camada plexiforme interna, onde funcionam como moduladores de sinal. Já foram introduzidas muitas classificações de células, incluindo uma que tem em conta o tamanho celular, e existem mais de 40 tipos diferentes de células amácrinas.
As células ganglionares são o elo de ligação entre a retina e o cérebro. O tipo mais comum de células ganglionares é o das células ditas anãs e parasol. O sinal luminoso, depois de percorrer as várias camadas da retina, acaba nas células ganglionares e é conduzida ao longo dos axónios destas células e do nervo óptico. O ponto na retina em que os axónios ganglionares se agregam para formar o nervo óptico é chamado o disco óptico. A maioria dos axónios termina no núcleo geniculado lateral, que trata de transmitir a informação recebida ao córtex visual. Algumas células ganglionares reagem a luz, mas visto que esta resposta é mais lenta do que a do cones e bastonetes, pensa-se que estará relacionada com a detecção de níveis luminosos ambientes e ajustamento do relógio biológico.
Processamento de sinal
editarTal como mencionado anteriormente, a retina é o principal componente sensorial do olho e contém todas as células sensíveis à luz. Sem esta componente central, o olho humano seria comparável a uma máquina fotográfica digital sem o sensor CCD (Charge Coupled Device). Esta secção pretende explicar como a percepção da luz é feita na retina, como o sinal óptico é transmitido para o cérebro e como o cérebro processa o sinal de forma a gerar informação suficiente para a tomada de decisões.
Criação dos sinais iniciais - Função fotorreceptora
editarA visão começa invariavelmente a estimulação de células fotorreceptoras por parte de raios visuais. Pigmentos visuais, uma variedade de enzimas e transmissores nos cones e bastonetes retinianos vão iniciar a conversão de ondas EM visíveis em potenciais eléctricos, um processo denominado transdução fotoeléctrica. Usando os bastonetes como exemplo: os raios de luz atingem as moléculas de rodopsina, moléculas transmembranares que se encontram nos discos dos bastonetes. cada molécula de rodopsina é constituída por um agregado de hélices chamado opsina, que envolve uma molécula de 11-cis retinal. Em moléculas biológicas, partes da molécula que vão provocar alterações conformacionais devidas a energia são chamadas de cromóforos. O 11-c retinal altera a sua conformação em resposta à energia luminosa, tornando-se em numa molécula de all-trans retinal, que separa as hélices da opsina, expondo sítios reactivos. Esta molécula de rodopsina "activada" é por vezes chamada metarodopsina II. A partir deste ponto, mesmo que o estímulo luminoso cesse, a reacção irá continuar. A metarodopsina II pode agora reagir com as cerca de 100 moléculas de uma Gs proteína chamada transducina, o que resulta em as depois do GDP ser convertido em GTP. A as-GTP activada depois liga-se à cGMP-phosphodiesterase(PDE), alterando as trocas iónicas, provocando uma fraca concentração citosólica de catiões e alterando consequentemente a potencial de membrana.
A transdução fotoeléctrica tem uma capacidade enorme de amplificação. Uma única molécula de retinal activada por um único quantum de luz provoca a hidrólise de até 106 moléculas de cGMP por segundo.
Fototransdução
editar- Um fotão de luz interage com retinal, num fotorreceptor. O retinal sobre isomerização, mudando de uma conformação 11-cis para all-trans.
- O retinal deixa de "caber" no sítio activa da opsina.
- A opsina sofre então uma alteração conformacional para metarhodopsina II.
- A metarhodopsina II é instável e subdivide-se, dando origem a opsina e all-trans retinal.
- A opsina activa a proteína reguladora transducina. Isto faz com que a transducina se dissocie do GDP ligado a ela e que se ligue a GDP. De seguida, a sunidade alfa dissocia-se da beta e da gamma, estando o GDP sempre ligado à subunidade alfa.
- O complexo subunidade alfa-GTP activa a fosfodiesterase (PDE).
- A PDE decompõe cGMP em 5'-GMP. Isto diminui a concentração de cGMP e, assim, os canais de sódio fecham.
- O fecho dos canais de sódio causa hiperpolarização da célula, devida à corrente de potássio.
- A hiperpolarização da célula causa o fecho dos canais de sódio sensíveis à voltagem.
- À medida que a concentração de cálcio dentro da célula fotoreceptora diminui, a quantidade do neurotransmissor glutamato que é libertado pela célula também diminui. Isto ocorre porque o cálcio é necessário para que as vesículas contendo glutamato se fusionem com a membrana celular e libertem os seu conteúdo.
- A diminuição da quantidade de glutamato libertado pelos fotoreceptores causa despolarização das células bipolares on center (associadas a cones e bastonetes) e hiperpolarização das células bipolares off (associadas a cones).
Sem a estimulação por ondas EM visíveis, os bastonetes contêm um cocktail de iões, proteínas e outras moléculas e possuem diferenças de potencial de membrana de cerca de -40 mV. Comparado com outras células neuronais (-65mV), este potencial é bastante alto. Neste estado, o neurotransmissor glutamato é libertado dos terminais axónicos e captado pelas células bipolares circundantes. Com a estimulação por onde EM visíveis e após a cascata anteriormente descrita se desenrolar, a diferença de potencial desce para -70 mV. Esta hiperpolarização causa uma redução na libertação de glutamato, afectando a activadade das células bipolares e os passos a jusante.
Processos similares existem nos cones e nas células ganglionares fotosenssíveis, mas utilizam opsinas diferentes. As fotopsinas I a III (amarelo-verde, verde e azul-violeta) são encontradas nos 3 diferentes tipos de cones e a melanopsina (azul), é encontrada nas células ganglionares fotossensíveis.
O Processamento de sinais na retina
editar
Diferentes células bipolares reagem de forma diferente às alterações da libertação de glutamato. As células bipolares ON despolarizam quando existem ondas EM visíveis e as célula ganglionar ON correspondente será activada. Por outro lado, as células bipolares OFF são hiperpolarizadas pela estimulação causada por ondas EM visíveis, a célula ganglionar OFF correspondente é inibida. Esta é a base da transmissão de sinal directo (direct signal flow). A transmissão lateral (lateral signal flow) começa nos bastonetes, atinge as células bipolares, as células amácrinas e as células bipolares OFF inibidas pelas células amácrinas associadas a bastonetes e as células bipolares ON serão estimuladas via uma sinapse eléctrica. Depois destes passos, o sinal chega às células ganglionares ON e OFF e toda a via lateral está estabelecida.
Quando o potencial de acção (PA) é ON, as células ganglionares são estimuladas pelas ondas EM visíveis. A frequência de PAs aumenta quando o potencial do sensor aumenta. Dito de outra forma, a frequência dos PAs depende da amplitude do potencial do sensor. A região das células ganglionares onde os efeitos estimulatórios e inibitórios influenciam a frequência dos PAs é chamado o campo receptivo (CR). Em relação às células ganglionares, os CR são geralmente compostos de duas regiões: uma zona central e uma zona periférica em forma de anel. Eles são distinguíveis durante a adaptação às ondas EM visíveis. Uma célula ganglionar com um centro ON e uma periferia OFF apresenta um aumento da frequência de PAs quando luz é incidida no centro e uma diminuição da frequência de PAs quando luz visível é incidida na periferia. As células ganglionares com um centro OFF e periferia ON apresentam o comportamento oposto. Os CR são organizados pelas células horizontais Esta estrutura dos CR permite realçar contrastes. Quando todo o CR é exposto a luz, a resposta da região central domina.
Transmissão de sinal ao córtex
editarComo mencionado anteriormente, os axónios das células ganglionares convergem para o disco óptico da retina, dando origem ao nervo óptico. As fibras da zona macular da retina ficam na porção central do nervo e as da metade temporal da retina permanecem à periferia. Uma decussação parcial das fibras ocorre quando o nervo deixa a cavidade orbitária. As fibras da metade nasal da retina decussam ao nível do chamado quiasma óptico e vão conectar-se com o córtex visual contralateral ao olho de onde provêm. Após o quiasma óptico, o nervo óptico passa a chamar-se tracto óptico, para assinalar uma nova estrutura do nervo. O objectivo da decussação é transmitir a porção direita do campo visual ao córtex visual esquerdo e vice-versa.

As fibras axonais das células ganglionares terminam no núcleo geniculado lateral (NGL), uma estrutura de grande importância no processamento de sinal visual. Cada NGL (existe um total de dois) está situado no tálamo e transmite informação ao córtex occipital ipsilateral. O NGL divide a informação transmitida pela retina ao cérebro em duas vias: a via parvocelular e a via magnocelular. A via parvocelular sinaliza cor e detalhe, enquanto que a via magnocelular detecta movimentos rápidos.

Os sinais de uma máquina fotográfica digital standard correspondem aproximadamente aos da via parvocelular. Para simular as respostas das vias parvocelulares, investigadores têm desenvolvido sistemas sensoriais neuromórficos, que tentam imitar a computação baseada em spikes dos sistemas neuronais biológicos. Este esquema de transmissão de sinal é chamado "address-event representation" (Liu and Delbruck 2010 [1]).
Em termos anatómicos, as células ganglionares ligadas às vias parvo- e magnocelulares projectam os seus axónios para as duas camadas magnocelulares ventrais e para as quatros parvocelulares dorsais do NGL. Cada uma das seis camadas do NGL recebe input do olho ipsi- ou contralateral. Por exemplo, algumas fibras ganglionares do olho esquerdo decussam e projectam para as camadas 1,4 e 6 no NGL direito enquanto que outras fibras ganglionares do olho direito se mantêm ipsilaterais e projectam para as camadas 2, 3 e 5. A partir do NGL, as informação de cada olho é separada.
Embora as diferentes metades do campo visual sejam processadas por hemisférios cerebrais diferentes, o campo visual é considerado uma unidade contínua e completa. Assim pensa-se que ambos os córtices occipitais estarão intimamente ligados, sobretudo através de uma estrutura chamada "corpo caloso".
Como dito anteriormente, as sinapses do tracto óptico são feitas ao longo das várias camadas do NGL. De seguida, os axónios destas células de terceira ordem estendem até à fissura calcarina de cada lobo occipital. Por causa do seu aspecto macroscópico, esta zona é chamada de córtex estriado, e corresponde ao nosso córtex visual primário, por vezes também chamado V1. Neste local, impulsos vindos dos dois olhos convergem nos mesmo neurónios corticais, o que permite usar este input para percepção e compreensão da imagem. O reconhecimento de padrões é uma função muito importante desta zona, sendo que lesões nesta área provocam problemas no reconhecimento visual e cegueira.
O processamento da informação da retina até ao cérebro é feita em point to point. Se toda a retina for estimulada iremos observar respostas em ambos os NGL e córtices occipitais. É possível traçar um mapeamento no córtex visual primário.
Qualquer detalhe adicional a esta via ultrapassa as intenções deste livro. Fique no entanto consciente de que existem muitos mais níveis e centros dedicados a tarefas específicas, como a cor, a orientação dos objectivos, frequências espaciais, emoções etc.
Processamento de informação no sistema visual
editarAgora que vimos os conceitos mais importantes no processamento de sinal no sistema visual, passemos para a compreensão ou percepção da informação sensorial processada. Chamamos percepção visual ao processo de tradução da informação recebida pelos olhos numa interpretação do estado externo das coisas. A percepção visual ajuda-nos a estar consciente do mundo e a compreendê-lo melhor. Com ela, aprendemos a reconhecer padrões, que depois utilizamos para tomar decisões. Dito de outra forma, a nossa sobrevivência depende da nossa percepção. A área da percepção visual foi dividida em várias subáreas, devido à sua complexidade e aos mecanismos especializados de cada. As sub-áreas incluem: percepção da cor, do movimento, de profundidade, reconhecimento de faces etc.
As hierarquias presentes no córtex visual primário
editar
Apesar do crescente poder computacional dos sistemas electrónicos, existem ainda muitas tarefas em que tanto animais como humanos apresentam uma performance altamente superior à dos computadores. Um exemplo será a percepção e contextualização de informação. O computador clássico (tanto o que tem no seu telemóvel como os super-computadores que ocupam salas inteiras) é um especialista em números: pode executar um número incrível de cálculos numa quantidade muito pequena de tempo. No entanto, falta-lhe a capacidade de criar abstracções da informação com a qual trabalha. Se ligar uma câmara ao seu computador, a imagem que o computador percebe é apenas uma matriz de pixels, uma tabela bidimensional de números. Em contrapartida, um humano seria capaz de reconhecer imediatamente a geometria da cena, os objectos na imagem e até talvez o contexto do que se estará a passar. Esta nossa capacidade é o resultado do trabalho de um sistema especializado: o sistema visual. Tudo o que vemos é processado de forma hierárquica, começando pelos aspectos mais simples de uma imagem até aos mais complexos e até à classificação de objectos em categorias. Assim, diz-se que o sistema visual possui uma hiearquia complexa. Esta hierarquia, presente em primatas, inspirou cientistas informáticos a criarem modelos de redes neuronais artificiais, que também possuem várias camadas, onde cada uma cria generalizações de ordem superior do input que recebe.
Cerca de metade do neocórtex visual humano é dedicado à visão. O processamento de informação visual ocorre em pelo menos 10 níveis funcionais. Os neurónios em áreas visuais mais primárias extraem características específicas da imagem em pequenas zonas do campo visual. À medida que a informação é transmitida para áreas de ordem superior, as representações começam a ser menos sensíveis a alterações no tamanho e na posição espacial dos objectos. Por outro lado, o CR dos neurónios destas áreas é maior, o que indica que estão à procura de características mais gerais da imagem. Esta estrutra hierárquica permite computações eficientes: áreas visuais mais avançadas podem usar a mesma informação de áreas mais abaixo na hierarquia. A descrição genérica fornecida por áreas primárias é usada por outras partes do cérebro para completar várias tarefas, como reconhecimento e categorização de objectos, manipulação de objectos, planeamento de movimentos etc.
Visão subcortical
editarO processamento neuronal da informação visual começa antes do sinal atingir estruturas corticais. Os fotorreceptoras da retina detectam luz e enviam sinais para as células ganglionares. O CR de uma fotorreceptor corresponde a um centésimo de grau (CR de um grau corresponde aproximadamente ao tamanho de uma polegar, se tiver o seu braço extendido à sua frente). Assim, a quantidade de inputs que uma célula ganglionar recebe e o tamanho do seu CR depende da sua localização (no centro da retina, uma célula ganglionar poderá receber informação de apenas cinco fotorreceptores, enquanto que na periferia o input pode vir de milhares de células). Isto implica uma maior resolução espacial no centro da retina, também chamado de fóvea. Devido a isto, os primatas possuem um mecanismo de controlo do olhar que os faz projectar a característica de interesse na fóvea.
As células ganglionares têm a capacidade de detectar vários aspectos de uma imagem: o contraste em termos de brilho e cor, a direcção e velocidade de movimentos. Todas estas características são a informação primária que é depois usada na pipeline de processamento de sinal. Se certos estímulos não forem detectados pelas células ganglionares, eles não serão apreendidos por outras estruturas a jusante.
As células ganglionares projectam para o NGL, que depois transmite a informação recebida para áreas corticais. Não existe nenhuma computação significativa no NGL, havendo quase uma correspondência one-to-one entre células ganglionares e células do NGL. No entanto, apenas 5% do input para o NGL provém da retina, o resto vindo de projecções corticais de feedback. Embora o sistema visual seja muitas vezes visto como um sistema feed-forward, as conexões laterais e de feedback são uma características proeminente deste sistema. A função das conexões recorrentes ainda não foi completamente estabelecida mas pensa-se que terão um papel a desempenhar em processos como atenção, imaginação e quando será necessário preencher o sinal com informação em falta.
Visão cortical
editar
O córtex visual pode ser dividido em três grandes partes: uma parte occipital que recebe o input fo NGL e projecta para as vias dorsal e ventral. A parte occipital inclui a áreas V1 a V4 e MT, que processam diferentes aspectos da informação visual e permitem uma representação genérica de uma cena visual. A via dorsal está envolvida na análise espacial e no planeamento de acções. A via ventral está envolvida no reconhecimento de objectos e na categorização destes.
A área V1 é a primeira área cortical a processar informação visual. É sensível a arestas, padrões lineares alternados, fins de linha, movimento, cor e disparidades (diferenças angulares entre projecções de um ponto nas retinas direita e esquerda). O exemplo mais claro de um processamento hierárquico e bottom-up é a combinação linear do input de várias células ganglionares para representar uma barra. Isto é feito pelas células simples de V1 e este processo foi primeiro descrito pelos prominentes neurocientistas Hubel e Wiesel. Este tipo de integração de informação implica que as células simples são sensíveis à localização exacta da barra e têm um CR relativamente pequeno. As células complexas de V1 recebem fibras das células simples e respondem a padrões lineares de forma independente à localização exacta destas. Estas células também possuem um CR maior, quando comparado com o das células simples.
A computação realizada neste passo poderá ser uma operação de tipo 'max', produzindo portanto respostas semelhantes, em termos de amplitude, ao estímulo mais intenso. Algumas células simples e complexas também conseguem detectar o fim de uma barra, e uma fracção das células de V1 são também sensíveis ao movimento local, dentro dos seus CR.
A área V2 consegue representar contornos mais complexos, incluindo textura, contornos ilusórios e contornos com border ownership. V2 também processa as disparidades detectadas em V1 e contém células sensíveis a disparidades relativas (a diferença entre as disparidades absolutas de dois pontos no espaço).
A área V4 recebe projecções de V2 e V3, mas pouco se sabe sobre a computação que terá lugar em V3. A área V4 contém neurónios sensíveis a contornos com diferentes curvaturas e a vértices com ângulos específicos. Uma outra função importante de V4 será a codificação de tonalidade que não varie com luminância. Esta forma de operar é diferente da que ocorre em V1, onde os neurónios reagem e respondem à oponência de cores ao longo dos dois eixos principais (vermelho-verde e amarelo-azul), e não às cores em si. A área V4 por sua vez projecta fibras para a via ventral, para o córtex temporal inferior (TI), que, através de estudos baseados em lesão, mostrou ser essencial para a discriminação de objectos.
Córtex temporal inferior (TI) - discriminação de objectos
editar
O córtex temporal inferior (TI) divide-se em duas áreas: TEO e TE. A área TEO integra informação sobre a forma e posição relativa de muitos elementos de contorno a contém sobretudo células que respondem a combinações visuais simples. O tamanho do CR dos neurónios da área TEO é de cerca 3 a 5 graus. A área TE tem células com CR significativamente maiores (10 a 20 graus) e que respondem a faces, mãos e complexas combinações de padrões. Estas células respondem a características que formam uma simples generalização do objecto de interesse (sendo no entanto estes padrões mais complexos do que meras barras ou manchas). Este facto foi mostrado usando o método de redução do estímulo de Tanaka et al., onde primeiro foi medida a resposta a um objecto, sendo este objecto depois substituído por representações progressivamente simplificadas (e isto até obter as características críticas que determinam a resposta dos neurónios de TE).
Parece que os neurónios de TI agregam várias características de complexidade média e de níveis inferiores na via ventral para construir modelos de partes de objectos. Os neurónios de TE que são selectivos a objectos específicos têm de cumprir dois pré-requisitos algo paradoxais: selectividade e invariabilidade. Têm de distinguir objectos usando a sua sensibilidade a certas características na imagem retiniana. No entanto, o mesmo objecto pode ser visto a partir de diferentes ângulos e distâncias, em condições de luminosidade muito díspares, o que leva à criação de imagens muito diferentes do mesmo objecto. Para tratar todas estas imagens como sendo equivalentes, as características invariantes têm de ser robustas contra certo tipo de transformações, como mudanças de posição, iluminação, tamanho da retina etc. Neurónios da área TE mostram invariabilidade a posição e também a oclusão parcial, posição em profundidade e direcção da iluminação. A rotação com profundidade mostrou ser a característica com menor invariância, à excepção da face humana.
As categorias de objectos ainda não estão explicitamente presentes na área TE: um neurónio pode responder a vários exemplares da mesma categoria (e.g. imagens de árvores), mas não responderá a todos. Por outro lado, também poderá responder a exemplares de diferentes categorias (e.g. árvores e não-árvores). O reconhecimento e a classificação de objectos muito provavelmente envolve a actividade conjunta de uma maior população de neurónios da área TE e a integração desta com o input de outras áreas cerebrais, como as responsáveis pela compreensão do contexto de uma cena. Foram recentemente feitas experiências que demonstraram que classificadores estatísticos (e.g. support vector machines) conseguem ser treinados para classificar objectos através da resposta de um pequeno número de neurónios TE. Assim, a população de neurónios TE consegue em princípio sinalizar categorias de objectos através da sua actividade conjunta. É interessante referir que existem relatos de neurónios altamente selectivos no córtex temporal medial que respondem a pistas muito específicas, como por exemplo a torre de Pisa em diferentes imagens ou à face de uma pessoa em particular.
A Aprendizagem no sistema visual
editarUm processo de aprendizagem pode alterar o padrão de selectividade neuronal, tendo como efeito que a aprendizagem se torna mais forte em níveis hierárquicos superiores. Não existe evidência de aprendizagem na retina. Para além disso, os mapas de orientação em V1 parecem, na sua maior parte, ser determinados geneticamente. No entanto, a prática de identificação de orientação melhora a codificação da orientação nos neurónios de V1, ao aumentar o declive da curva de afinação. Efeitos similares foram observados na área V4. Na área TE, pequenas quantidades de treino visual têm notáveis efeitos fisiológicos na percepção visual, tanto a nível unicelular como a nível mascroscópico (visto em fMRI). Por exemplo, misturar dois objectos aumenta a semelhança percebida. De forma geral, parece que até o córtex visual do adulto apresenta uma considerável plasticidade, e o nível de plasticidade pode ser aumentado de forma significativa (por exemplo, administrando fármacos específicos ou vivendo num ambiente rico).
Redes neuronais profundas
editarTal como na hierarquia do córtex visual primário, arquitecturas neuronais profundas tentam modelar abstracções de alto nível do input fornecido recorrendo a múltiplos níveis de transformações não-lineares. O modelo proposto por Hubel e Wiesel (onde a informação é integrada e propagada numa cascata, partindo da retina, atingindo o NGL, as células simples e complexas de V1) inspirou a criação de um dos primeiros modelos de arquitecturas de deep learning: o neocognitron, um modelo de rede neuronal com múltiplas camadas. Foi usada para diferentes tarefas de reconhecimento de padrão, incluindo o reconhecimento de dígitos escritos à mão. No entanto, era necessário muito tempo para treinar a rede neuronal (na ordem dos dias), e desde a sua criação nos anos 80, a área de deep learning não recebeu muita atenção; isto até aos anos 2000, com a abundância de dados digitais e a invenção de algoritmos de treino mais rápidos. As redes neuronais profundas provaram ser muito boas em tarefas que, até há relativamente pouco tempo, só os humanos pareciam ser capazes de resolver. Exemplos serão o reconhecimento da face de pessoas específicas em fotografias, a compreensão do discurso humano (até certo ponto) e a tradução de línguas estrangeiras. Por outro lado, provaram ser muito úteis na indústria e em investigação (ajudam a procurar potenciais fármacos candidatas, a mapear redes neuronais biológicas e a prever a função de proteínas). É necessário notar as arquitecturas de deep learning são muito superficialmente inspiradas em redes neuronais biológicas e constituem muito mais um feito da área da informática / machine learning do que da das neurociências. O paralelo básico que é traçado é que as redes neuronais artificias são compostas por unidades (neurónios) que integram a informação de forma não-linear e projectam sinais entre si. Finalmente as redes neuronais artificias contêm diferentes níveis de representação abstrata dos dados. O algoritmo de aprendizagem e as descrições matemáticas dos "neurónios" usados em deep learning são muito diferentes dos processos biológicos que ocorrem no cérebro. Assim, a investigação em deep learning, embora catalise muitos avanços na área da inteligência artificial, apenas fornece insights limitados à neurociência.
Fontes
editar- Artigos sobre as hierarquias profundas do sistema visual
- Kruger, N.; Janssen, P.; Kalkan, S.; Lappe, M.; Leonardis, A.; Piater, J.; Rodriguez-Sanchez, A. J.; Wiskott, L. (August 2013). "Deep Hierarchies in the Primate Visual Cortex: What Can We Learn for Computer Vision?". IEEE Transactions on Pattern Analysis and Machine Intelligence 35 (8): 1847–1871. doi:10.1109/TPAMI.2012.272.
- Poggio, Tomaso; Riesenhuber, Maximilian (1 November 1999). Nature Neuroscience 2 (11): 1019–1025. doi:doi:10.1038/14819.
- A experiência de redução de estímulo
- Tanaka, Keiji (March 1996). "Inferotemporal Cortex and Object Vision". Annual Review of Neuroscience 19 (1): 109–139. doi:10.1146/annurev.ne.19.030196.000545
- Evidência de aprendizagem no sistema visual
- Li, Nuo; DiCarlo, James J. (23 September 2010). "Unsupervised Natural Visual Experience Rapidly Reshapes Size-Invariant Object Representation in Inferior Temporal Cortex". Neuron 67 (6): 1062–1075. doi:10.1016/j.neuron.2010.08.029.
- Raiguel, S.; Vogels, R.; Mysore, S. G.; Orban, G. A. (14 June 2006). "Learning to See the Difference Specifically Alters the Most Informative V4 Neurons". Journal of Neuroscience 26 (24): 6589–6602. doi:10.1523/JNEUROSCI.0457-06.2006.
- Schoups, A; Vogels, R; Qian, N; Orban, G (2 August 2001). "Practising orientation identification improves orientation coding in V1 neurons.". Nature 412 (6846): 549-53. PMID 11484056.
- Uma review recente e acessível do estado da investigação em deep learning
- Jones, Nicola (8 January 2014). "Computer science: The learning machines". Nature 505 (7482): 146–148. doi:10.1038/505146a.
Percepção de movimento
editarA percepção de movimento é o processo de inferência da velocidade e direcção de objectos em movimento. As áreas V5 (em humanos) e MT (medial temporal) em primatas são as responsáveis pela percepção cortical de movimento. A área V5 integra o córtex extrastriado, que corresponde ao córtex anatomicamente próximo do córtex visual primário. A função da área V5 é uma de detectar a velocidade e direcção dos estímulos visuais e de integrar os sinais de movimento visual local em movimento global. O córtex visual primário situa-se no lobo occipital de ambos os hemisférios cerebrais. Esta área representa a primeira etapa de processamento cortical da informação visual e contém uma representação completa do campo visual. A diferença entre V5 e V1 (outro nome para o córtex visual primário) é que V5 é capaz de integrar o movimento de sinais locais ou partes individuais de um objecto de forma a representar o movimento do objecto por inteiro. A área V1, por sua vez, responde a moção local que ocorre dentro de CRs e as estimativas de muitos neurónios são integradas na área V5.
O movimento é definido como definido como uma alteração da iluminação retiniana ao longo do espaço e do tempo. Os sinais de movimento são classificados em "movimentos de primeira ordem" e "movimentos de segunda ordem". Vamos de seguida descrever estes tipos de movimento.

A "percepção de movimento de primeira ordem" refere-se ao movimento percebido quando dois ou mais estímulos visuais aparecem e desaparecem ao longo do tempo e produzem percepções de movimento diferentes. Moção de primeira ordem também é chamada "moção aparente", e é usada na televisão e em filmes. Um exemplo é o "movimento beta", uma ilusão em que imagens fixas parecem mexer-se. Estas imagens dão a impressão de movimento porque se alteram mais rapidamente do que limiar de detecção do olho. O nervo óptico humano responde a alterações luminosas a cada dez ciclos por segundo, portanto qualquer alteração mais rápida será registada com um movimento em contínuo, e não como imagens separadas.
A "moção de segunda ordem" refere-se a moção que ocorre quando um contorno em movimento é definido por contraste, textura, cintilação ou outra qualidade que não resulta num aumento de luminância ou energia de moção da imagem. Existe evidência que sugere o processamento inicial de moção de primeira e segunda ordens é feito por vias distintas. Os mecanismos de segunda ordem possuem uma resolução temporal inferior e não são muito específicos em termos das frequências espaciais às quais respondem. Moção de segunda ordem produz um after effect inferior. Tanto os sinais de primeira como de segunda ordem são combinados na área V5.
Neste capítulo, analisamos os conceitos de percepção de movimento e análise de movimento. Por outro lado, explicamos a razão pela qual estes termos não devem ser interpretados como sendo sinónimos. Iremos analisar os mecanismos através dos quais o movimento é percebido (e.g. sensores de movimentos e feature tracking). Existem três principais modelos teóricos que tentam descrever a função dos neurónios sensores de movimento. Foram conduzidas experiências para confirmar a validez destes modelos. Infelizmente, os resultados não são conclusivos, e parece que nenhum modelo único consegue descrever o funcionamento dos sensores de movimento. Contudo, cada um dos modelos simula diferentes aspectos dos sensores. Algumas destas propriedades serão descritas. Finalmente, este capítulo irá apresentar algumas ilusões de movimento, que demonstram que o nosso sentido de movimento pode ser enganado por factores externos estáticos que estimulam os sensores de movimento da mesma maneira que a verdadeira moção.
Análise de movimento e percepção de movimento
editarOs conceitos de análise de movimento e percepção de movimento são muitas vezes erradamente usados como se se tratassem de sinónimos. Ambos os conceitos estão de facto interligados, mas não são a mesma coisa.
A "análise de movimento" refere-se ao mecanismo através do qual sinais de movimento são processados. Da mesma maneira que a percepção de movimento não depende necessariamente de sinais gerados por moção de imagens na retina, a análise de moção pode ou não levar à percepção de movimento. Um exemplo disto será a vecção, que ocorre quando uma pessoa sente movimento quando está de facto estática, mas o objecto que observa está em movimento. O fenómeno de vecção mostra que um objecto pode ser analizado, mesmo que a percepção de movimento não venha do objecto em si. Esta definição de análise de moção sugere que movimento é uma propriedade fundamentalmente da imagem. No campo visual, este é analisado em todos os pontos. Os resultados desta análise são usados para derivar informação perceptual.
A "percepção de movimento" refere-se ao processo de aquisição de conhecimento perceptual sobre o movimento de objectos e superfícies numa imagem. A moção é percebida ou graças a sensores locais na retina a feature tracking. Os sensores de movimento local são neurónios especializados sensíveis a moção e análogos a sensores especializados de cor. Feature tracking é uma forma indirecta de detectar movimento, e consiste na inferência de movimento a partir de alterações da posição retiniana de objectos ao longo do tempo.
Sensores de movimento
editarA detecção de movimento é o primeiro estádio do processamento visual, e acontece graças a processos neuronais especializados que respondem a informação que transmita alterações locais de intensidade da imagem ao longo do tempo.
A moção é percebida de forma independente de outras propriedades de imagens, seja qual for o local onde incide. A existência de sensores de movimento já foi provada, e estes operam de forma local, em todos os pontos de uma imagem. Os sensores de movimento são sensores neuronais especializados, localizados na retina e capazes de detectar movimentos produzido por dois curtos e pequenos flashes de luz que estão tão próximos um dos outro que não poderiam ser detectados por feature tracking. Existem três modelos que tentam descrever a forma como estes sensores operam. Estes modelos são independentes uns dos outros e tentam modelas características específicas da percepção de movimento. Embora não haja evidência suficiente a favor de um modelo e em detrimento dos outros dois, estes modelos sempre representam certas funções destes sensores.

O detector de Reichardt
O detector de Reichardt é usado para modelar a forma como sensores de movimento respondem a sinais de moção de primeira ordem. Quando um objecto se move de ponto A a B no campo visual, dois sinais são gerados: um antes do movimento começar e outro após o movimento ter sido completado. Este modelo detecta esse movimento ao perceber alterações na luminância de um ponto na retina e correlacionando-as com alterações na luminosidade detectadas noutro ponto próximo após um curto espaço de tempo. o detector de Reichardt opera baseado no princípio de correlação (relação estatística que envolve dependência). Este modelo interpreta o sinal de moção através de uma correlação espaciotemporal de luminância detectada em pontos próximos. O modelo usa o facto de que dois CRs colocados a diferentes pontos da trajectória de um objecto em movimento recebem o mesmo sinal, mas em tempos diferentes. O modelo detector de Reichardt contém dois detectores espacialmente próximos um do outro. O output dos detectores é multiplicado (correlacionado): um sinal é multiplicado por um segundo sinal que nada mais é do que uma versão mais tardia do mesmo sinal. O mesmo procedimento é repetido mas na direcção inversa à do movimento (o sinal mais tardio torna-se no primeiro sinal vice versa). De seguida, a diferença entre estas duas multiplicações é calculada, e o resultado indica a velocidade do movimento. A resposta do detector depende da fase do estímulo, do contraste e da velocidade. Muitos detectores afinados para diferentes velocidades são necessários de maneira a codificar a verdadeira velocidade de um padrão. A maior evidência para este tipo de detector vem de estudos de discriminação de direcção em alvos quase não perceptíveis.
Motion-Energy Filtering
Um filtro de motion energy é um modelo de sensores de movimento baseado no princípio de filtros invariante em fase. Este modelo constrói filtros espaciotemporais orientados no espaço e no tempo, de forma a detectar a estrutura de padrões em movimento. Consiste de filtros separáveis, no qual os perfis espaciais mantêm a mesma forma mas são alterados pelo valor dos filtros temporais. Estes filtros estabelecem uma correspondência com padrões em movimento ao adicionar filtros separáveis. para cada direcção de movimento, dois filtros espaciais são gerados: um simétrico (tipo barra) e outro assimétrico (tipo aresta). A soma dos quadrados destes filtros é chamada a energia de moção. A diferença de sinal entre as duas direcções é chamada a energia oponente. Este resultado é de seguida dividido por output de outro filtro ao quadrado, filtro esse que detecta o contraste estático. Esta divisão é feita de forma a ter em atenção o efeito do contraste no movimento. Os filtros de motion energy conseguem modelar vários fenómenos de movimento, but produz uma medida independente de fase, que aumenta com a velocidade, mas não indica um valor fidedigno de velocidade.
Gradientes espaciotemporais

Este modelo de sensores de movimento foi originalmente desenvolvido na área da visão de computadores e é baseado no princípio de que o rácio da derivada temporal do brilho de uma image e da derivada espacial do brilho da imagem dá a velocidade do movimento. É importante referir que, em picos e valas de uma imagem, este modelo não irá fornecer uma resposta adequada, visto que a derivada será zero. De maneira a resolver este problema, as derivadas de primeira ordem e de ordem mais elevada em relação ao espaço e ao tempo têm de ser analisadas. Os gradientes espaciotemporais constituem um bom modelo para determinar a velocidade de um movimento em todos os pontos de uma imagem.
Sensores de movimentos são selectivos em relação à orentiação
editarUma das propriedade dos sensores de movimento é sua selectividade face a diferentes orientações, o que reduz a análise de movimento a um problem unidimensional. De facto, estes sensores só conseguem registar movimento numa dimensão ao longo de um eixo ortogonal à orientação preferida do sensor. Um estímulo que contenha padrões com uma única orientação só pode ser visto em movimento se avançar numa direcção ortogonal à orientação do estímulo. Sinais unidimensionais de movimento fornecem informação ambígua em relação ao movimento de objectos bidimensionais. Uma segunda fase de análise de movimento é necessária de forma a discernir a verdadeira direcção de movimento de um objecto 2-D ou padrão. Sinais de moção 1-D de sensores selectivos para diferentes orientações são combinados de forma a produzir um sinal de movimento 2-D não ambíguo. A análise de moção bidimensional depende dos sinais de sensores altamente selectivos e pouco selectivos.
Feature Tracking
editarUma outra forma de detectar movimento é através de feature tracking. Feature Tracking consiste em verificar se as características locais de um objecto mudaram de posição e em inferir movimento se esta mudança ocorrer. Nesta secção, iremos mencionar uns aspectos de feature tracking. Os chamados feature trackers falham quando um estímulo em movimento ocorre muito rapidamente. Estes têm a vantagem, em relação aos detectores do movimento, de que conseguem detectar o movimento de um objecto mesmo se o movimento for separado por intervalos intermitentes sem movimento. Os feature trackers conseguem separar estes dois estádios. Os sensores de movimento, por outro lado, integrariam os intervalos sem movimento com a imagem do estímulo em movimento e veriam um movimento contínuo. Os feature trackers operam baseados na localização das características identificadas. Por esta razão, eles possuem um limiar mínimo de detecção que corresponde à precisão com a qual a localizações das características podem ser discriminadas. Não mostram efeitos a posteriori de moção(motion aftereffects), que são ilusões visuais que resultam de um processo de adaptação visual. Estas ilusões ocorrem quando, após termos observado um objecto em movimento, um objecto imóvel nos é apresentado e parece estar a mover-se na direcção contrária à do estímulo prévio. É impossível para este mecanismo monitorizar múltiplos movimentos em diferentes partes do campo visual e ao mesmo tempo. Em contrapartida, vários tipos de movimentos não são um problema para os sensores de movimento, visto que operam em paralelo ao longo de todo o campo visual
Experiências usando a informação mencionada anteriormente foram conduzidas e traçaram conclusões interessantes sobre feature trackers. Experiências com estímulos breves mostraram que padrões de cor e de contraste com alto contraste não são detectados pelos feature trackers e sim pelos sensores de movimento. Experiências com intervalos sem movimento foram conduzidas e confirmaram que o fenómeno de feature tracking pode ocorrer co =m intervalos sem movimento no ecrã. Apenas com contraste elevado que os sensores de movimento detectam o movimento de estímulos cromáticos e padrões de contraste. Com baixos contrastes, os feature trackers analisam tanto a moção de padrões cromáticos como de contraste, enquanto que, com contrastes altos, os sensores de moção analisam os padrões de contraste. Experiências nas quais os sujeitos fazem múltiplas avaliações de movimento sugerem que o processo de feature tracking ocorre sob controlo consciente e que representa a única forma de analisar a moção de envelopes de contraste in displays de baixo contraste. Estes resultados são consistentes com a noção de que o movimento destes dois tipos de padrões depende de feature tracking, excepto quando as cores se situam bem acima do limiar ou o contraste médio é alto. A grande conclusão destas experiências é que é provavelmente o processo de feature tracking que permite a percepção de padrões de contraste e cromáticos.
Ilusões de movimento
editarDada a forma como a percepção de movimento funciona, algumas imagens estáticas poderão parecer estar em movimento. Estas imagens dão-nos dicas sobre as premissas que o sistema visual faz, e são chamadas ilusões visuais.
Uma famosa ilusão visual relacionada com sinais de moção de primeira ordem é o fenómeno de Phi, uma ilusão óptica que nos faz perceber movimento em vez de que uma sequência de imagens. Esta ilusão permite-nos experienciar filmes como algo contínuo e não como imagens separadas. O fenómeno de Phi faz-nos ver uma sequência de imagens estáticas que são trocadas a um ritmo constante como um aparente movimento constante. O fenómeno de Phi não deve ser confundido com o movimento Beta, visto que este é um movimento aparente causado por impulsos luminosos apresentados em sequência, enquanto que o primeiro é um movimento aparente provocado por impulsos luminosos estáticos.
Ilusões de movimentos acontecem quando a percepção de movimentos, a análise de moção e a interpretação destes sinais são erróneos, deixando que o nosso sistema visual crie uma ilusão de movimento. Estas ilusões são classificadas de acordo com o processo que permite que elas aconteçam. De facto, são classificadas em ilusões relacionadas com a percepção de movimento, com a integração 2D e com a interpretação 3D.
As ilusões mais populares relacionadas com percepção de movimento são a moção a quatro tempos, RDKs e ilusões de sinais de moção de segunda ordem. As ilusões relacionadas com a integração 2D mais conhecidas são as de captura de movimento, movimento de padrões em xadrez e repulsão directa. Finalmente, as ilusões relacionadas com a interpretação 3D são as de moção transformacional, profundidade cinética, movimento de sombra, biológico, stereocinético, movimento de figura implícito e movimento a dois tempo. Existem muito mais ilusões de movimento, e todas demonstram algo interessante sobre os mecanismos humanos de detecção, percepção e análise de movimento. Para mais informações, não hesite em consultar o link seguinte: http://www.lifesci.sussex.ac.uk/home/George_Mather/Motion/
Problemas em aberto
editarEmbora não entendamos todos os detalhes relacionados com a percepção de movimento, compreender os mecanismos através dos quais o movimento é percebido tal como entender as ilusões de movimento permite já dar uma boa revisão do estado da arte neste campo. Alguns problemas relacionados com a percepção de movimento ainda em aberto são os mecanismos de formação de imagens 3D em moção global e o problema da apertura. Os sinais globais de movimento oriundos da retina são integrados de forma a obter um sinal 2-D do sinal do moção global. No entanto, ainda não se sabe como a moção global 3D é formada. O problema da apertura ocorre porque cada CR dentro do sistema visual cobre apenas uma pequena porção do mundo visual, o que leva a ambiguidades na percepção. O problema da apertura refere-se à questão de que um contorno em movimento, quando observado localmente, é consistente com diferentes possibilidades de moção. Esta ambiguidade é de origem geométrica (um movimento paralelo ao contorno não pode ser detectado, visto que alterações neste componente de moção não alteram a imagem, quando observada através da apertura). O único componente que pode ser medido é a velocidade ortogonal à orientação do contorno. Por esta razão, a velocidade do movimento poderá ser qualquer uma dentro da família de movimentos ao longo de uma linha no espaço de velocidades. O problema da apertura é não só observado em contorno rectos mas também em contornos curvados, visto que são aproximadamente rectos quando observados localmente. Embora os mecanismos que permitem de resolver o problema da apertura são desconhecidos, existem algumas hipóteses sobre como pode ser resolvido. Por exemplo, seria possível resolver este problema combinando a informação ao longo do espaço e de diferentes contornos do objecto.
Conclusões
editarNeste capítulo, introduzimos a noção de percepção de movimento e os mecanismos pelos quais o nosso sistema visual detecta movimento. Ilusões de movimento mostram como sinais de moção podem ser enganadores e levar a conclusões incorrectas sobre moção. É importante lembrar que a percepção e a análise de movimento não são o mesmo conceito. Sensores de movimento e feature trackers complementam-se de forma a permitir ao nosso sistema visual detectar movimento.
A percepção de movimento é um fenómeno complexo, e uma área de investigação com muitos problemas em aberto. Este capítulo pretendeu descrever modelos de funcionamento dos sensores de moção e hipóteses sobre as características de feature trackers. No entanto, mais experiências são necessárias para aprender mais sobre as características destes mecanismos e sermos capazes de construir modelos que simulem de forma mais precisa os processos biológicos que ocorrem no nosso sistema visual
A variedade de mecanismos de análise e percepção de moção descrita neste capítulo, tal como a sofisticação dos modelos artificiais desenhados para os descrever demonstram bem a complexidade que existe na forma como o nosso córtex processa sinais do mundo externo. Milhares de neurónios especializados integram e interpretam porções de sinais locais para formar imagens globais de objectos em movimento. Ao entender a quantidade de actores e processos que precisam de trabalhar em conjunto para nós humanos percebermos movimentos, percebemos o quão incrível esta nossa capacidade é, sobretudo tendo em conta que o fazemos com uma aparente e quase desconcertante facilidade.
Percepção de cor
editarIntrodução
editarOs humanos (juntamente com outros primatas como os macacos e os gorilas) são os mamíferios com melhor percepção de cor [1] . Assim, não é uma coincidência que a cor desempenhe um papel importante numa variedade de aspectos. Por exemplo, a cor é útil para discriminar e diferenciar objectos, superfícies, cenas naturais e até faces [2],[3]. Color is also an important tool for nonverbal communication, including that of emotion [4].
Durante muitas décadas, mostrar as ligações entre as propriedades físicas da cor e as suas qualidades perceptuais tem sido um desafio. Tipicamente, são estudadas recorrendo a duas abordagens diferentes: analisando a resposta comportamental causada por cor (numa área chamada "psicofísica") e a resposta fisiológica causada pela detecção de cor[5].
Aqui só nos iremos focar neste último aspecto. O estudo da base fisiológica da visão cromática, sobre a qual quase nada se sabia até à segunda metade do século vinte, tem avançado de forma menos rápida mas segura desde os anos 1950. Tem havido bastante progresso em muitas áreas, sobretudo ao nível da análise dos receptores. Graças aos métodos da biologia molecular, foi possível revelar detalhes até lá desconhecidos sobre a base genética dos pigmentos dos cones. Por outro lado, tem-se mostrado cada vez mais áreas corticais que são influenciadas pelos estímulos visuais, embora a correlação entre a percepção cromática com reacções fisiológicas ligadas ao comprimento de onda e que ocorram fora dos fotorreceptores não seja tão fácil de discernir.[6].
Neste capítulo iremos tentar explicar os aspectos básicos dos diferentes processos envolvidos na percepção de cor ao longo da via visual, desde a retina até ao córtex visual. Para obter mais detalhes anatómicos, confira a secção sobre anatomia do sistema visual deste Wikilivro.
Percepção cromática na retina
editarTodas as cores que podem ser discriminadas pelos humanos podem ser produzidas por uma mistura de apenas três cores primárias (básicas). Inspirado por esta ideia de mistura de cores, tem sido proposto que a cor é detectada por três classes de sensores, cada um tenho uma sensibilidade máxima a uma parte diferente do espectro de luz visível [1]. Em 1853 foi proposto pela primeira vez e de forma explícita que três graus de liberdade dentro da combinação normal de cores[7]. Isto foi depois confirmado em 1886 [8] (with remarkably close results to recent studies [9], [10]).
Estes sensores de cores propostos são na realidade os cones acima mencionados (Nota: neste capítulo, iremos apenas falar de cones. Os bastonetes contribuem para a visão apenas em níveis baixos de intensidade de luz). Apesar de se saber que têm um efeito na percepção de cores, a sua influência é muito pequena e não será portanto abordada aqui.)[11].
Os cones correspondem a um dos dois tipos de fotorreceptores presentes na retina, estando particularmente concentrados na fóvea. A tabela abaixo lista os três tipos de cones. Estes podem ser distinguidos pelos diferentes tipos de pigmento de rodopsina que apresentam. As curvas de absorção correspondentes são representadas na curva abaixo.
| Nome | Maior sensibilidade cromática | Pico de absorção [nm] |
|---|---|---|
| S, SWS, B | Azul | 420 |
| M, MWS, G | Verde | 530 |
| L, LWS, R | Vermelho | 560 |
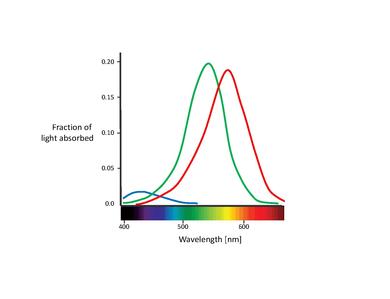
Embora não se tenha atingido nenhum consenso sobre como nomear os diferentes tipos de cones, as designações mais comummente usadas referecem-se ou ao pico de absorção espectral ou à cor à qual são sensíveis (vermelho, verde, azul)[6]. Neste texto, iremos recorrer à designação S-M-L (short, medium, and long wavelength), visto que estes nomes são adequadamente descritivos. A nomenclatura azul-verde-vermelho é algo enganosa, visto que todos os tipos de cones são sensíveis a um grande intervalo de comprimentos de onda.
Uma característica importante destes cones é a sua distribuição relativa na retina. Sabe-se hoje que o cones S estão presentes apenas em baixas concentrações ao longo da retina, estando completamente ausentes na porção mais central da fóvea. Aliás, estão de tal forma dispersos que se pensa que não desempenham um papel importante na visão espacial, embora sejam capazes de mediar uma fraca percepção de contornos[12]. A fóvea é dominada pelos cones L e M. A proporção de ambos é tipicamente apresentada sob a forma de um rácio. Diferentes valores foram reportados para o rácio L/M, indo de 0.67 [13] até 2 [14], sendo o último mais comummente aceite. A razão pela qual existem sempre mais cones L do que M não é clara. É interessante notar que o rácio de cones L e M per se não tem quase nenhum impacto na visão. Este facto mostra claramente a plasticidade do nosso cérebro, sendo capaz de interpretar os vários sinais que podem ser gerados pelos cones [15], [16].
É também importante notar a a sobreposição dos espectros de absorção dos cones L e M. Enquanto o espectro dos cones S está claramente separado dos outros, os picos dos espectros de absorção dos cones L e M estão a 30 nm de distância um do outro, com forte sobreposição das curvas. Existe portanto uma grande correlação na resposta destas duas classes de cones. Isto é explicado pelo facto de que, para atingir a maior acuidade visual possível ao nível da fóvea, o sistema visual trata os cones L e M de forma igual, não tenho em conta os seus espectros de absorção. Assim, qualquer tipo de diferença leva a uma deterioração do sinal de luminância [17]. Dito de outra forma, a pequena separação entre cones L e M pode ser interpretada como um compromisso entre a necessidade de visão cromática de alto contraste e a de grande acuidade visual em termos de luminância. Esta ideia é congruente com a falta de cones S na porção central da fóvea, onde a acuidade visual é máxima. Por outro lado, as semelhanças entre a os espectros de absorção dos cones L e M também podem ser explicados pela sua origem genética. Assume que ambos os tipos de cones evoluíram recentemente (há cerca de 35 milhões de ano), a partir de um "antepassado" comum, enquanto que se pensa que os cones S se terão separado do "antepassado" muito antes[11].
Os diferentes espectros de absorção dos cones são a marca da visão cromática humana. Esta teoria resolveu um problema muito antigo: embora possamos ver milhões de cores diferentes (os seres humanos conseguem distinguir 7 a 10 milhões de cores diferentes)[5], as nossas retinas simplesmente não têm espaço suficiente para acomodar um detector individual para cada cor em todos os pontos da retina.
Da retina ao cérebro
editarOs sinais que são transmitidos da retina a níveis superiores não são meras representações ponto por ponto dos sinais oriundos dos receptores, mas antes consistem de combinações sofisticadas dos sinais dos receptores. O objectivo desta secção é providenciar uma breve descrição das vias que este tipo de informação segue.
Uma vez que a imagem óptica da retina sofre transdução em sinais químicos e eléctricos dentro dos fotorreceptores, os sinais modelados pela amplitude são convertidos em representações moduladas pela frequência ao nível das células ganglionares e a níveis superiores. Nestas células neuronais, a magnitude do sinal é representado em termos do número de spikes por segundo, em vez da diferença de potencial ao nível da membrana. De maneira a explicar e representar as propriedades destas células, é muito útil usar o conceito de campos receptivos (CR).
Um CR é uma representação gráfica de uma área do campo visual à qual uma dada célula responde. Por outro lado, a natureza da resposta é tipicamente indicada pelas várias regiões do CR. Por exemplo, podemos considerar o CR de uma célula fotorreceptora como sendo uma pequena área circular que representa o tamanho e a localização da sensibilidade daquele fotorreceptor particular dentro do campo visual. A figura abaixo mostra exemplos de CR de células ganglionares que tipicamente apresentam um antagonismo centro/periferia. O CR à esquerda na figura ilustra uma resposta central positiva (tem um centro "ON"). Este tipo de resposta é tipicamente gerado por um input positivo de um cone único, rodeado por uma resposta negativa gerada pelos vários cones à volta. Assim, a resposta desta célula ganglionar será composta pelos inputs dos vários cones com sinais positivos e negativos. Desta forma, a célula não só responde aos pontos de luz como também serve de um detector de cantos (ou mais, correctamente, de manchas).
O CR à direita na figura ilustra uma resposta central negativa (possui um centro dito "OFF"). Tipicamente, células de centro "ON" e "OFF" irão ocorrer na mesma localização espacial e receberão input dos mesmo fotorreceptores.
A figura abaixo mostra que, para além dos antagonismo espacial, as células ganglionares também podem apresentar uma oponência espectral. Por exemplo, a parte esquerda da figura inferior ilustra a oponência vermelho-verde, com o centro a receber um input positivo de um cone L e a periferia a receber um input negativo dos cones M. Por outro lado, a parte direita da figura inferior mostra a versão "OFF" desta célula. Assim, antes da informação visual ter sequer deixado a retina, já houve alguma forma de processamento, com efeitos profundos sobre a aparência das cores. Existem outros tipos e variedades de células ganglionares, mas todas partilham o mesmo conceito.


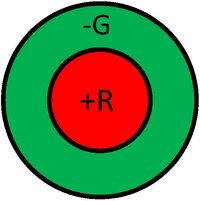

A caminho do córtex visual primário, os axónios das células ganglionares reunem-se para formar o nervo óptico, que projecta fibras para o núcleo geniculado lateral (NGL), no tálamo. A codificação de informação no nervo óptico é altamente eficiente e mantém o número de fibras necessárias num mínimo (limitado pelo tamanho do nervo óptico). Aqui também se pretende manter o ponto cega retiniano (largura de aproximadamente 5° e altura de cerca de 7°) o mais pequeno possível.
Além do mais, as células ganglionares apresentadas não apresentariam resposta quando expostas a iluminação uniforme, visto que as áreas positivas e negativas se balanceiam. Dito de outra forma, os sinais transmitidos não se correlacionam. Por exemplo, a informação de pontos próximos numa cena natural apresentação uma grande correlação espacial e portanto são altamente previsíveis [18]. A inibição lateral entre células vizinhas no interior da retina ajuda a minimizar esta correlação espacial, aumentando assim a eficiência do sistema. Este processo pode ser interpretado como servindo um propósito de compressão da imagem, ainda na retina.
Dada a sobreposição dos espectros de absorção dos cones L e M, os seus sinais também acabam por estar altamente correlacionados. Neste caso, a eficiência da computação é melhorada combinando os sinais dos cones de maneira a minimizar a dita correlação. É possível entender este processo usando uma técnica chamada Principal Component Analysis (PCA). A PCA é um método estatístico usado para reduzir a dimensionalidade de um dado grupo de variáveis ao transformar as variáveis originais num set de novas variáveis: os componentes principais (CP). O primeiro CP tem em conta a quantidade máxima de variância total nas variáveis originais, enquanto que o segundo CP engloba a quantidade máxima de variância que não foi tida em conta pelo primeiro componente, e assim de seguida. Para além do mais, os CP são linearmente independentes e ortogonais no espaço dos parâmetros. A maior vantagem da PCA é que apenas alguns dos CP mais fortes são suficientes para cobrir a vasta maioria da variabilidade do sistema [19]. Este esquema tem sido usado com as funções de absorção dos cones [20] e mesmo com espectros que ocorrem de forma natural[21],[22]. Os CP que foram encontrados no espaço de excitação dos cones produzida por objectos naturais são 1) um eixo de luminância onde os cones L e M são adicionados (L+M), 2) a diferença do sinais L e M (L-M) e 3) um eixo de cor onde o sinal do cone S é diferenciado da soma L+M. Estes canais, derivados de uma abordagem matemática/computacional, coincide com os três canais retino-geniculados descobertos em experiências electrofisiológicas [23],[24]. Usando estes mecanismos, a informação redundante é eliminada da retina.
Existem três canais de informação que de facto comunicam esta informação da retina às células ganglionares e ao NGL. Estes canais são diferentes não apenas em termos das suas propriedades cromáticas, mas também em termos do seu substrato anatómico. Estes canais colocam importantes limitações em tarefas de cor básicas, tais como detecção e discriminação.
No primeiro canal, o output dos cones L e M é transmitido de forma sinergética a células bipolares difusas e de seguida às camadas magnocelulares (M) do NGL (a não confundir com os cones M da retina)[24]. Os CR da células M são compostas de um centro e de uma zona periférica, que funcionam como antagonistas espaciais. As células M possuem uma sensibilidade e alto contraste para estímulos luminosos, mas não mostram nenhuma resposta a combinações de inputs L-M oponentes[25]. No entanto, porque os pontos nulos da diferentes células M variam ligeiramente, a resposta populacional nunca é zero. Esta propriedade é na realidade passada a áreas corticais com um input predominante de células M[26].
A via parvocelular (P) inicia-se nos outputs individuais dos cones L ou M e nas células ganglionares anãs. Este oferecem depois input a células P retinianas[11]. Na fóvea, os CR das células P são formados por cones L ou M únicos. A estrutura da periferia do CR de uma célula P é ainda hoje debatida. Contudo, a teoria mais aceite diz que a periferia consiste de um tipo de cone específico, resultando num CR espacialmente oponente para estímulos luminosos[27]. Parvocellular layers contribute with about 80 % of the total projections from the retina to the LGN[28].
Finalmente, a via coniocelular (K), recentemente descoberta, leva consigo a maior dos sinais oriundos dos cones S[29]. Grupos deste tipo de cones projectam para células bipolares especiais, que por sua vez providenciam input a pequenas células ganglionares específicas. Estas não são usualmente oponentes em termos espaciais. Os axónios das pequenas células ganglionares projectam-se para finas camadas do NGL (adjacentes às camadas parvocellulares)[30].
Embora as células ganglionares terminem no NGL (estabelecendo sinapses com as células do NGL), parece haver uma correspondência individual entre células ganglionares e células do NGL. O NGL parece funcionar como uma estação que retransmite os sinais que recebe. No entanto, provavelmente terá também uma função visual, visto que existem projecções neuronais do córtex de volta ao NGL que poderão servir de mecanismo de feedback. Os axónios das células do NGL projectam para V1, no córtex visual do lobo occipital.
Percepção de cor no cérebro
editarNo córtexa, as projecções das vias magno-, parvo- e coniocelulares terminam em diferentes camadas do córtex visual primário. As fibras magnocelulares inervam principalmente as camadas 4Cα e 6. Os neurónios parvocelulares projectam sobretudo para 4Cβ e para as camadas 4A e 6. Os neurónios coniocelulares terminam nos blobs ricos em citocromo oxidase das camadas 1, 2 e 3[31].
Uma vez no córtex visual, a codificação da informação visual torna-se significativamente mais complexa. Da mesma maneira, os outputs de vários fotorreceptores são combinados e comparados para produzir a resposta das células ganglionares, os outputs das várias células do NGL são combinados e comparados para produzir uma resposta cortical. À medida que os sinais ascendem a cadeia de processamento de sinal cortical, este processo repte-se com um nível crescente de complexidade, até ao ponto em que os CR começam a perder o seu significado. Contudo, algumas funções e processos foram identificados e estudados em regiões específicas do córtex visual.
Na região V1 (córtex estriado), neurónios duplamente oponente (neurónios cujo CR com regiões ON/OFF cromaticamente e espacialmente opostas) comparam os sinais de cor ao longo do espaço visual[32]. Estas células consituem 5 a 10% das células de V1. O seu aspecto grosseiro e pequena concentração combinam com a fraca resolução espacial, em termos de visão cromática [1]. Por outro lado, elas não são sensíveis à direcção de movimento dos estímulos (ao contrário de alguns neurónios em V1) e, assim, dificilmente irão contribuit para a percepção de movimento[33]. Contudo, dada a natureza especializada dos seus CR, este tipo de células constituem a base neuronal para efeitos de contraste cromático e um meio eficiente de codificar cor[34],[35]. Outras células de V1 respondem a outros tipos de estímulo, tal como cantos orientados, várias frequências espaciais e temporais, em particular localizações espaciais, e a uma combinação destas características, entre outros. Adicionalmente, é possível encontrar células que combinam o input das células do NGL de forma linear e outras que efectuam esta combinação de forma não-linear. Estas respostas são necessárias para suportar capacidades visuais avançadas, tais como cor.

Existe substancialmente menos informação sobre as propriedades cromáticos dos neurónios da área V2 em comparação com V1. À primeira vista, não parece haver grandes diferenças em termos de codificação da cor em V1 e em V2[36]. Uma excepção é o surgimento de uma nova classe de células cromáticas complexas[37]. Assim, já foi sugerido que V2 está envolvido na elaboração de cor. Contudo, isto é ainda ponto algo controverso, que ainda não foi confirmado.
Seguindo o conceito modular desenvolvido após a descoberta da dominância ocular funcional em V1, e considerando a segregação anatómica entre as vias P, M e K (descritas na secção 3), tem sido sugerido que um sistema especializado dentro do córtex visual, dedicado à análise de cor, existe[38]. V4 é a região que historicamente tem atraído mais atenção como possível "área da cor" no cérebro. Isto é por causa de um estudo importante que afirmou que V4 contém 100% das células sensíveis à tonalidade[39]. No entanto, esta afirmação foi discutida num número considerável de estudos subsequentes, alguns reportando que apenas 16% dos neurónios em V4 são sensíveis à cor[40]. Actualmente, o conceito mais aceite é que V4 contribui não só para a percepção de cor, mas também para a perceção de forma, para a atenção visual e para a estereopsia. Estudos recentes têm-se focado noutras regiões cerebrais, tentando encontrar a "área cerebral da cor". A TEO tem sido por exemplo considerada[41] and PITd[42]. A relações destas regiões entre si ainda são muito debatidas. Para reconciliar as várias partes envolvidas no debate, alguns usam o termo córtex temporal ínfero-posterior (PIT) para designar uma região que inclui V4, TEO e PITd[1].
Se as respostas corticias em células de V1, V2 e V4 já são por si uma tarefa complicada, o nível de complexidade das respostas visuais complexas de cerca de 30 zonas visuais é enorme. A figura 4 mostrou uma pequena porção da conectividade das diferentes áreas corticais que foram identificadas[43].
A este ponto, torna-se cada vez mais difícil explicar a função de células corticais únicas em termos simples. De facto, a função de uma célula única pode nem sequer conter significado, visto que a representação de várias percepções tem de ser distribuída ao longo de grupos de células em vários pontos do córtex.
Mecanismos de adaptação na visão cromática
editarEmbora os investigadores tenham tentado explicar o processamento de sinais cromáticos no sistema visual humano, é importante entender que a percepção de cor não é um processo fixe. De facto, existe uma variedade de mecanismos dinâmicos que servem para optimizar a resposta visual de acordo com o ambiente observado. Os mecanismos de adaptação ao escuro, à luz e à cor são particularmente importantes.
Adaptação ao escuro
editarA adaptação ao escuro refere-se à alteração da sensibilidade visual que ocorre quando o nível de iluminação diminui. A resposta do sistema visual a uma iluminação reduzida é ficar mais sensível, aumentado a sua capacidade de produzir uma resposta visual adequada mesmo quando as condições de iluminação são subóptimas[44].
A figura 5 mostra a recuperação da sensibilidade visual após a exposição a um ambiente de muita iluminação e passagem para um ambiente escuro[43]. Primeiro, os cones vão ficando progressivamente mais sensíveis, até a curva estabilizar após alguns minutos. Depois, após aproximadamente 10 minutos, a sensibilidade visual é quase constante. Nesta altura, o sistema de bastonetes, que tem um tempo de recuperação de maior, já recuperou sensibilidade suficiente para dominar os cones e portanto assumir o controlo da sensibilidade total. A sensibilidade dos bastonetes também melhora progressivamente, até apresentar um comportamento assintótico após cerca de 30 minutos. Dito de outra forma, os cones são responsáveis pela recuperação da sensibilidade durante os primeiros 10 minutos. Depois disto, os bastonetes assumem o controlo da sensibilidade e atingem a sua sensibilidade após cerca de 30 minutos.
Este é apenas um entre vários mecanismos neuronais que permitem uma boa adaptação a condições de fraca iluminação. Outro mecanismos neuronais, incluindo o famosos reflexo pupilar e a deplecção ou regeneração de fotopigmento assumem maior importância nas células retinianas e noutros mecanismos de nível mais elevado, incluindo interpretações cognitivas.
Adaptação à luz
editarA adaptação à luz é essencialmente o processo contrário à adaptação ao escuro. De facto, os mecanismos fisiológicos por detrás deste fenómeno são os mesmo em ambos os processos. Contudo, é importante considerá-los em separado, visto que as suas propriedades visuais diferem.

A adaptação à luz ocorre quando os níveis de iluminação são aumentados. Assim, o sistema visual tem de ficar menos sensível de forma a produzir percepções úteis, visto que há mais luz visível disponível. O sistema visual tem um intervalo de output dinâmico limitado para os sinais que produzem as nossas percepções. Contudo, o mundo externo contém níveis de iluminação cobrindo pela menos mais 10 ordens de grandeza. Felizmente, nós raramente precisamos de ver ao mesmo tempo todo o intervalo de níveis de iluminação.
Em níveis de alta luminosidade, a adaptação é conseguida através de "branqueamento" dos fotopigmentos. Isto aumenta a captura de fotões nos receptores e protege a resposta dos cones, impedindo saturação em ambientes com fundos brilhantes. O mecanismo de adaptação à luz ocorre primariamente na retina[45]. De facto, alterações de ganho são largamente específicas a cones e a adaptação concentra os sinais em áreas que não serão maiores de o diâmetro de um cone individual[46],[47]. Isto aponta para uma localização da adaptação à luz que pode começar logo ao nível dos receptores. Contudo, parece haver mais do que um ponto de aumento ou diminuição da sensibilidade. Alguns aumentos são extremamente rápidos, enquanto que outros demorar segundos ou até minutos a estabilizar[48]. Usually, light adaptation takes around 5 minutes (six times faster than dark adaptation). Isto poderá apontar para a influência de sítios pós-receptivos.
A figura 6 mostra exemplos de adaptação à luz[43]. Se usássemos apenas uma função de resposta para mapear o largo intervalo de intensidades no output do sistema visual, então teríamos um pequeno intervalo à disposição para uma dada cena. Torna-se claro que, com tal função de resposta, o contraste percebido em qualquer cena seria limitado e a sensibilidade visual a alterações severamente degradada devido a problemas relacionados com o rácio sinal para ruído. Este é o caso que a linha tracejada pretende ilustrar. Por outro lado, as linhas sólidas representam famílias de respostas. Estas curvas mapeiam o intervalo de iluminação útil numa cena num intervalo completo a dinâmico de output visual. Assim, obtemos a melhor percepção possível para cada situação. O processo de adaptação à luz pode ser visto como uma forma de deslizar a curva de resposta visual ao longo do nível de iluminação, até o nível óptimo para um dado conjunto de iluminação ser atingido.
Adaptação cromática
editarO conceito geral de adaptação cromática consiste na variação da altura das três curvas de resposta espectral cónica. Este ajustamento acontece porque a adaptação à luz ocorre de forma independente dentro de cada classe de cone. Uma formulação específica desta hipótese é conhecida como a "adaptação de von Kries". Esta hipótese diz que a resposta adaptativa ocorre em cada um dos três tipos de cones de forma separada e que é o equivalente a multiplicar as suas sensibilidades espectrais fixas por uma constante[49]. Se o coeficientes (também conhecidos "como coeficientes de von Kries") são inversamente proporcionais à absorção de luz de casa tipo de cone (isto é, uma menor absorção vai requerer um coeficiente maior), então este método mantém uma resposta média constante por classe de cone. Isto oferece um mecanismo simples mais poderoso para manter a percepção de cor do objectos, apesar das alterações em termos de iluminação. Cumprindo um certo número de condições, este método fornece uma boa aproximação dos efeitos de adaptação à luz sobre a sensibilidade à e aparência de cor[50],[51].
A forma mais fácil de imaginar a adaptação cromática é examinando um objecto branco sob diferentes condições de iluminação. Por exemplo, consideremos que estamos a examinar uma folha de papel sob luz do dia, luz fluorescente e sob iluminação incandescente. A luz natural contém relativamente muito menos energia de curto comprimento de onda que a luz fluorescente, e a iluminação incandescente contém comparativamente muito mais energia de longo comprimento de onda que a luz fluorescente. Contudo, apesar das diferentes condições de iluminação, o papel parece reter, pelo menos em parte, o seu aspecto branco, sob os três diferentes tipos de iluminação. Isto acontece porque o sistema de cones S se torna relativamente menos sensível quando exposto a luz natural (de forma a compensar a energia adicional de pequeno comprimento de onda), enquanto que os cones L se tornam comparativamente menos sensíveis sob luz incandescente (de forma a compensar a energia adicional de grande comprimento de onda)[43].
Bibliografia
editar- Conway, Bevil R (2009). "Color vision, cones, and color-coding in the cortex". The neuroscientist 15: 274-290.
- Russell, Richard and Sinha, Pawan} (2007). "Real-world face recognition: The importance of surface reflectance properties". Perception 36 (9).
- Gegenfurtner, Karl R and Rieger, Jochem (2000). "Sensory and cognitive contributions of color to the recognition of natural scenes". Current Biology 10 (13): 805-808.
- Changizi, Mark A and Zhang, Qiong and Shimojo, Shinsuke (2006). "Bare skin, blood and the evolution of primate colour vision". Biology letters 2 (2): 217-221.
- Beretta, Giordano (2000). Understanding Color. Hewlett-Packard.
- Boynton, Robert M (1988). "Color vision". Annual review of psychology 39 (1): 69-100.
- Grassmann, Hermann (1853). "Zur theorie der farbenmischung". Annalen der Physik 165 (5): 69-84.
- Konig, Arthur and Dieterici, Conrad (1886). "Die Grundempfindungen und ihre intensitats-Vertheilung im Spectrum". Koniglich Preussischen Akademie der Wissenschaften.
- Smith, Vivianne C and Pokorny, Joel (1975). "Spectral sensitivity of the foveal cone photopigments between 400 and 500 nm". Vision research 15 (2): 161-171.
- Vos, JJ and Walraven, PL (1971). "On the derivation of the foveal receptor primaries". Vision Research 11 (8): 799-818.
- Gegenfurtner, Karl R and Kiper, Daniel C (2003). "Color vision". Neuroscience 26 (1): 181.
- Kaiser, Peter K and Boynton, Robert M (1985). "Role of the blue mechanism in wavelength discrimination". Vision research 125 (4): 523-529.
- Paulus, Walter and Kroger-Paulus, Angelika (1983). "A new concept of retinal colour coding". Vision research 23 (5): 529-540.
- Nerger, Janice L and Cicerone, Carol M (1992). "The ratio of L cones to M cones in the human parafoveal retina". Vision research 32 (5): 879-888.
- Neitz, Jay and Carroll, Joseph and Yamauchi, Yasuki and Neitz, Maureen and Williams, David R (2002). "Color perception is mediated by a plastic neural mechanism that is adjustable in adults". Neuron 35 (4): 783-792.
- Jacobs, Gerald H and Williams, Gary A and Cahill, Hugh and Nathans, Jeremy (2007). "Emergence of novel color vision in mice engineered to express a human cone photopigment". Science 315 (5819): 1723-1725.
- Osorio, D and Ruderman, DL and Cronin, TW (1998). "Estimation of errors in luminance signals encoded by primate retina resulting from sampling of natural images with red and green cones". JOSA A 15 (1): 16-22.
- Kersten, Daniel (1987). "Predictability and redundancy of natural images". JOSA A 4 (112): 2395-2400.
- Jolliffe, I. T. (2002). Principal Component Analysis. Springer.
- Buchsbaum, Gershon and Gottschalk, A (1983). "Trichromacy, opponent colours coding and optimum colour information transmission in the retina". Proceedings of the Royal society of London. Series B. Biological sciences 220 (1218): 89-113.
- Zaidi, Qasim (1997). "Decorrelation of L-and M-cone signals". JOSA A 14 (12): 3430-3431.
- Ruderman, Daniel L and Cronin, Thomas W and Chiao, Chuan-Chin (1998). "Statistics of cone responses to natural images: Implications for visual coding". JOSA A 15 (8): 2036-2045.
- Lee, BB and Martin, PR and Valberg, A (1998). "The physiological basis of heterochromatic flicker photometry demonstrated in the ganglion cells of the macaque retina". The Journal of Physiology 404 (1): 323-347.
- Derrington, Andrew M and Krauskopf, John and Lennie, Peter (1984). "Chromatic mechanisms in lateral geniculate nucleus of macaque". The Journal of Physiology 357 (1): 241-265.
- Shapley, Robert (1990). "Visual sensitivity and parallel retinocortical channels". Annual review of psychology 41 (1): 635--658.
- Dobkins, Karen R and Thiele, Alex and Albright, Thomas D (2000). "Comparison of red--green equiluminance points in humans and macaques: evidence for different L: M cone ratios between species". JOSA A 17 (3): 545-556.
- Martin, Paul R and Lee, Barry B and White, Andrew JR and Solomon, Samuel G and Ruttiger, Lukas (2001). "Chromatic sensitivity of ganglion cells in the peripheral primate retina". Nature 410 (6831): 933-936.
- Perry, VH and Oehler, R and Cowey, A (1984). "Retinal ganglion cells that project to the dorsal lateral geniculate nucleus in the macaque monkey". Neuroscience 12 (4): 1101--1123.
- Casagrande, VA (1994). "A third parallel visual pathway to primate area V1". Trends in neurosciences 17 (7): 305-310.
- Hendry, Stewart HC and Reid, R Clay (2000). "The koniocellular pathway in primate vision". Annual review of neuroscience 23 (1): 127-153.
- Callaway, Edward M (1998). "Local circuits in primary visual cortex of the macaque monkey". Annual review of neuroscience 21 (1): 47-74.
- Conway, Bevil R (2001). "Spatial structure of cone inputs to color cells in alert macaque primary visual cortex (V-1)". The Journal of Neuroscience 21 (8): 2768-2783.
- Horwitz, Gregory D and Albright, Thomas D (2005). "Paucity of chromatic linear motion detectors in macaque V1". Journal of Vision 5 (6).
- Danilova, Marina V and Mollon, JD (2006). "The comparison of spatially separated colours". Vision research 46 (6): 823-836.
- Wachtler, Thomas and Sejnowski, Terrence J and Albright, Thomas D (2003). "Representation of color stimuli in awake macaque primary visual cortex". Neuron 37 (4): 681-691.
- Solomon, Samuel G and Lennie, Peter (2005). "Chromatic gain controls in visual cortical neurons". The Journal of neuroscience 25 (19): 4779-4792.
- Hubel, David H (1995). Eye, brain, and vision. Scientific American Library/Scientific American Books.
- Livingstone, Margaret S and Hubel, David H (1987). "Psychophysical evidence for separate channels for the perception of form, color, movement, and depth". The Journal of Neuroscience 7 (11): 3416-3468.
- Zeki, Semir M (1973). "Colour coding in rhesus monkey prestriate cortex". Brain research 53 (2): 422-427.
- Conway, Bevil R and Tsao, Doris Y (2006). "Color architecture in alert macaque cortex revealed by fMRI". Cerebral Cortex 16 (11): 1604-1613.
- Tootell, Roger BH and Nelissen, Koen and Vanduffel, Wim and Orban, Guy A (2004). "Search for color 'center(s)'in macaque visual cortex". Cerebral Cortex 14 (4): 353-363.
- Conway, Bevil R and Moeller, Sebastian and Tsao, Doris Y (2007). "Specialized color modules in macaque extrastriate cortex". 560--573 56 (3): 560-573.
- Fairchild, Mark D (2013). Color appearance models. John Wiley & Sons.
- Webster, Michael A (1996). "Human colour perception and its adaptation". Network: Computation in Neural Systems 7 (4): 587 - 634.
- Shapley, Robert and Enroth-Cugell, Christina (1984). "Visual adaptation and retinal gain controls". Progress in retinal research 3: 263-346.
- Chaparro, A and Stromeyer III, CF and Chen, G and Kronauer, RE (1995). "Human cones appear to adapt at low light levels: Measurements on the red-green detection mechanism". Vision Research 35 (22): 3103-3118.
- Macleod, Donald IA and Williams, David R and Makous, Walter (1992). "A visual nonlinearity fed by single cones". Vision research 32 (2): 347-363.
- Hayhoe, Mary (1991). Adaptation mechanisms in color and brightness. Springer.
- MacAdam, DAvid L (1970). Sources of Color Science. MIT Press.
- Webster, Michael A and Mollon, JD (1995). "Colour constancy influenced by contrast adaptation". Nature 373 (6516): 694-698.
- Brainard, David H and Wandell, Brian A (1992). "Asymmetric color matching: how color appearance depends on the illuminant". JOSA A 9 (9): 1443-1448.
- ↑ 1,0 1,1 1,2 1,3 Conway, Bevil R (2009). "Color vision, cones, and color-coding in the cortex". The neuroscientist 15: 274-290.
- ↑ Russell, Richard and Sinha, Pawan} (2007). "Real-world face recognition: The importance of surface reflectance properties". Perception 36 (9).
- ↑ Gegenfurtner, Karl R and Rieger, Jochem (2000). "Sensory and cognitive contributions of color to the recognition of natural scenes". Current Biology 10 (13): 805-808.
- ↑ Changizi, Mark A and Zhang, Qiong and Shimojo, Shinsuke (2006). "Bare skin, blood and the evolution of primate colour vision". Biology letters 2 (2): 217-221.
- ↑ 5,0 5,1 Beretta, Giordano (2000). Understanding Color. Hewlett-Packard.
- ↑ 6,0 6,1 Boynton, Robert M (1988). "Color vision". Annual review of psychology 39 (1): 69-100.
- ↑ Grassmann, Hermann (1853). "Zur theorie der farbenmischung". Annalen der Physik 165 (5): 69-84.
- ↑ Konig, Arthur and Dieterici, Conrad (1886). "Die Grundempfindungen und ihre intensitats-Vertheilung im Spectrum". Koniglich Preussischen Akademie der Wissenschaften.
- ↑ Smith, Vivianne C and Pokorny, Joel (1975). "Spectral sensitivity of the foveal cone photopigments between 400 and 500 nm". Vision research 15 (2): 161-171.
- ↑ Vos, JJ and Walraven, PL (1971). "On the derivation of the foveal receptor primaries". Vision Research 11 (8): 799-818.
- ↑ 11,0 11,1 11,2 Gegenfurtner, Karl R and Kiper, Daniel C (2003). "Color vision". Neuroscience 26 (1): 181.
- ↑ Kaiser, Peter K and Boynton, Robert M (1985). "Role of the blue mechanism in wavelength discrimination". Vision research 125 (4): 523-529.
- ↑ Paulus, Walter and Kroger-Paulus, Angelika (1983). "A new concept of retinal colour coding". Vision research 23 (5): 529-540.
- ↑ Nerger, Janice L and Cicerone, Carol M (1992). "The ratio of L cones to M cones in the human parafoveal retina". Vision research 32 (5): 879-888.
- ↑ Neitz, Jay and Carroll, Joseph and Yamauchi, Yasuki and Neitz, Maureen and Williams, David R (2002). "Color perception is mediated by a plastic neural mechanism that is adjustable in adults". Neuron 35 (4): 783-792.
- ↑ Jacobs, Gerald H and Williams, Gary A and Cahill, Hugh and Nathans, Jeremy (2007). "Emergence of novel color vision in mice engineered to express a human cone photopigment". Science 315 (5819): 1723-1725.
- ↑ Osorio, D and Ruderman, DL and Cronin, TW (1998). "Estimation of errors in luminance signals encoded by primate retina resulting from sampling of natural images with red and green cones". JOSA A 15 (1): 16-22.
- ↑ Kersten, Daniel (1987). "Predictability and redundancy of natural images". JOSA A 4 (112): 2395-2400.
- ↑ Jolliffe, I. T. (2002). Principal Component Analysis. Springer.
- ↑ Buchsbaum, Gershon and Gottschalk, A (1983). "Trichromacy, opponent colours coding and optimum colour information transmission in the retina". Proceedings of the Royal society of London. Series B. Biological sciences 220 (1218): 89-113.
- ↑ Zaidi, Qasim (1997). "Decorrelation of L-and M-cone signals". JOSA A 14 (12): 3430-3431.
- ↑ Ruderman, Daniel L and Cronin, Thomas W and Chiao, Chuan-Chin (1998). "Statistics of cone responses to natural images: Implications for visual coding". JOSA A 15 (8): 2036-2045.
- ↑ Lee, BB and Martin, PR and Valberg, A (1998). "The physiological basis of heterochromatic flicker photometry demonstrated in the ganglion cells of the macaque retina". The Journal of Physiology 404 (1): 323-347.
- ↑ 24,0 24,1 Derrington, Andrew M and Krauskopf, John and Lennie, Peter (1984). "Chromatic mechanisms in lateral geniculate nucleus of macaque". The Journal of Physiology 357 (1): 241-265.
- ↑ Shapley, Robert (1990). "Visual sensitivity and parallel retinocortical channels". Annual review of psychology 41 (1): 635--658.
- ↑ Dobkins, Karen R and Thiele, Alex and Albright, Thomas D (2000). "Comparison of red--green equiluminance points in humans and macaques: evidence for different L: M cone ratios between species". JOSA A 17 (3): 545-556.
- ↑ Martin, Paul R and Lee, Barry B and White, Andrew JR and Solomon, Samuel G and Ruttiger, Lukas (2001). "Chromatic sensitivity of ganglion cells in the peripheral primate retina". Nature 410 (6831): 933-936.
- ↑ Perry, VH and Oehler, R and Cowey, A (1984). "Retinal ganglion cells that project to the dorsal lateral geniculate nucleus in the macaque monkey". Neuroscience 12 (4): 1101--1123.
- ↑ Casagrande, VA (1994). "A third parallel visual pathway to primate area V1". Trends in neurosciences 17 (7): 305-310.
- ↑ Hendry, Stewart HC and Reid, R Clay (2000). "The koniocellular pathway in primate vision". Annual review of neuroscience 23 (1): 127-153.
- ↑ Callaway, Edward M (1998). "Local circuits in primary visual cortex of the macaque monkey". Annual review of neuroscience 21 (1): 47-74.
- ↑ Conway, Bevil R (2001). "Spatial structure of cone inputs to color cells in alert macaque primary visual cortex (V-1)". The Journal of Neuroscience 21 (8): 2768-2783.
- ↑ Horwitz, Gregory D and Albright, Thomas D (2005). "Paucity of chromatic linear motion detectors in macaque V1". Journal of Vision 5 (6).
- ↑ Danilova, Marina V and Mollon, JD (2006). "The comparison of spatially separated colours". Vision research 46 (6): 823-836.
- ↑ Wachtler, Thomas and Sejnowski, Terrence J and Albright, Thomas D (2003). "Representation of color stimuli in awake macaque primary visual cortex". Neuron 37 (4): 681-691.
- ↑ Solomon, Samuel G and Lennie, Peter (2005). "Chromatic gain controls in visual cortical neurons". The Journal of neuroscience 25 (19): 4779-4792.
- ↑ Hubel, David H (1995). Eye, brain, and vision. Scientific American Library/Scientific American Books.
- ↑ Livingstone, Margaret S and Hubel, David H (1987). "Psychophysical evidence for separate channels for the perception of form, color, movement, and depth". The Journal of Neuroscience 7 (11): 3416-3468.
- ↑ Zeki, Semir M (1973). "Colour coding in rhesus monkey prestriate cortex". Brain research 53 (2): 422-427.
- ↑ Conway, Bevil R and Tsao, Doris Y (2006). "Color architecture in alert macaque cortex revealed by fMRI". Cerebral Cortex 16 (11): 1604-1613.
- ↑ Tootell, Roger BH and Nelissen, Koen and Vanduffel, Wim and Orban, Guy A (2004). "Search for color 'center(s)'in macaque visual cortex". Cerebral Cortex 14 (4): 353-363.
- ↑ Conway, Bevil R and Moeller, Sebastian and Tsao, Doris Y (2007). "Specialized color modules in macaque extrastriate cortex". 560--573 56 (3): 560-573.
- ↑ 43,0 43,1 43,2 43,3 Fairchild, Mark D (2013). Color appearance models. John Wiley & Sons.
- ↑ Webster, Michael A (1996). "Human colour perception and its adaptation". Network: Computation in Neural Systems 7 (4): 587 - 634.
- ↑ Shapley, Robert and Enroth-Cugell, Christina (1984). "Visual adaptation and retinal gain controls". Progress in retinal research 3: 263-346.
- ↑ Chaparro, A and Stromeyer III, CF and Chen, G and Kronauer, RE (1995). "Human cones appear to adapt at low light levels: Measurements on the red-green detection mechanism". Vision Research 35 (22): 3103-3118.
- ↑ Macleod, Donald IA and Williams, David R and Makous, Walter (1992). "A visual nonlinearity fed by single cones". Vision research 32 (2): 347-363.
- ↑ Hayhoe, Mary (1991). Adaptation mechanisms in color and brightness. Springer.
- ↑ MacAdam, DAvid L (1970). Sources of Color Science. MIT Press.
- ↑ Webster, Michael A and Mollon, JD (1995). "Colour constancy influenced by contrast adaptation". Nature 373 (6516): 694-698.
- ↑ Brainard, David H and Wandell, Brian A (1992). "Asymmetric color matching: how color appearance depends on the illuminant". JOSA A 9 (9): 1443-1448.